Abstract
Mycobacterium ulcerans infection causes Buruli ulcer, a chronic and destructive necrotizing skin ulcer in humans. The symptoms of Buruli ulcer are mainly caused by unique toxic macrolides, named mycolactone. The suppression of Th1-type immune responses and inflammatory responses is caused by mycolactone in humans and animal models. However, the effects of mycolactone on serum immune responses remain unclear. In this study, we administered model antigens with partially purified mycolactone into mice to examine its effect on antibody production in serum. Mycolactone-containing fraction suppressed antibody production against co-administered antigens in a dose-dependent manner. The suppressive effect was not observed when the antigens and mycolactone preparation were injected into different sites. Additionally, the effect was demonstrated only against co-administered antigens. Moreover, mycolactone-containing fraction was considered safe even at doses ten times higher than the dose that suppressed the antibody responses, suggesting potential usefulness of mycolactone as a new immunoregulatory agent to specifically prevent antibody response against co-administered antigens.
Keyword
Mycobacterium ulcerans, Buruli ulcer, immunosuppression, mycolactone
Introduction
Buruli ulcer is a necrotizing skin infection caused by Mycobacterium ulcerans [1-3]. The disease mainly prevails in West Africa, and its incidence is increasing in Asia, Australia, and Latin America [1-3]. The pathogenesis of Buruli ulcer depends on mycolactone, a lipid toxin produced by M. ulcerans [4]. Mycolactone is a polyketide-derived macrolide [5] that exerts cytopathic effects on cultured L929 murine fibroblasts [4,6]. Injection of mycolactone into guinea pig produced an ulcer similar to Buruli ulcer [4]. Mycolactone inhibits the production of various cytokines, chemokines, and other secreted immune modulators [7] from immune cells such as monocytes [8], macrophages [9], dendritic cells [10], and T cells [11-13]. When a mycolactone-deficient strain of M. ulcerans was injected into model animals, a granuloma was successfully developed and the bacteria were cleared from the infected regions [14,15], suggesting that mycolactone has a suppressive effect on cell-mediated immune responses. However, the effects of mycolactone on serum immune responses remain poorly understood. We previously demonstrated that mice immunized with formalin-killed whole-cells of M. ulcerans did not produce M. ulcerans-specific serum IgG [16]. On the other hand, mice immunized with dewaxed whole-cells produced high-level of titers of M. ulcerans-specific IgG [16], leading us to hypothesize that waxy cell walls, containing mycolactone, inhibited the antibody response. In this study, we examined the inhibitory effect of mycolactone on the antibody production against co-administered antigens.
Materials and methods
Mice
Specific-pathogen-free female BALB/c mice were obtained from Japan SLC (Hamamatsu, Japan). All mice were housed in an animal care facility at Kitasato University. Mice were allowed free access to water and a standard diet with a 12 h light/dark cycle, at a room temperature of 23 ± 2°C with 55 ± 10% humidity. All cages were filled with wood shavings. The study was approved by the Animal research Committee of Kitasato University and conducted according to the guidelines of the Ministry of Education, Culture, Sports, Science, and Technology of Japan.
Bacterial strains and culture conditions
M. ulcerans TMC1615 (a clinical isolate producing mycolactone A/B) and M. ulcerans ATCC19423 (a mycolactone-deficient strain) were used in this study. We confirmed that mycolactone activity was not detected in a lipid fraction prepared from M. ulcerans ATCC19423 by the method described below. For routine propagation, bacteria were grown in Middlebrook 7H9 broth (BD Biosciences, Sparks, MD, USA) supplemented with 0.05% (w/v) Tween 80 and 10% (v/v) Middlebrook OADC enrichment (BD Biosciences) at 32°C for three weeks.
Partially purified mycolactone preparations
The preparation of acetone soluble lipid (ASL), a partially purified mycolactone, was performed as described by George et al. [6]. In brief, the bacterial culture was centrifuged to collect cells. A mixture of chloroform and methanol (2:1, v/v) was added to the cell pellet and stirred gently for 4 h at room temperature. After centrifugation (23,800 × g, 10 min, 4°C), the organic solvent layer was transferred into a brown glass bottle and the solvent was removed by using an evaporator. Ice-cold acetone was added to dissolve lipids, except phospholipids. After centrifugation (950 × g, 30 min, 4°C), the supernatant was transferred into a brown glass bottle and the solvent was removed by using an evaporator. Acetone was added and the solution, which we refer to as acetone soluble lipids (ASL), was stored at -20°C until further use.
Cytopathicity assay
CCL-1 mouse fibroblast, a L929 cell derivative, was purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). The cells were grown in Eagle’s minimum essential medium containing 10% heat-inactivated fetal calf serum (FCS, Moregate Biotech, Bulimba, QLD, Australia), penicillin (100 U/mL), and streptomycin (100 µg/mL) at 37°C in an atmosphere of 5% CO2 in air. Serial two-fold dilutions of test samples were added to each well of a 96-well microtiter plate, which contained 5 × 104 cells, and incubated for 48 h at 37°C. Then, 10 µL of AlamarBlue (Life Technologies, Carlsbad, CA, USA) was added to each well and incubated for 2 h. Optical density at 560 nm (OD560) and OD595 of the contents of each well were measured with a microplate reader. A Metabolic activity Index (MI) was calculated by using the following formula: MI = [(OD560-OD595)sample – (OD560-OD595)blank]/[(OD560-OD595)control – (OD560-OD595)blank]. Treatment with mycolactone-containing lipids led to the rounding up of CCL-1 cells [4, 6] and decreased MI value. A strong correlation was observed between percentages of cells with morphological change (cytopathic activity unit, CPU) [6] and MI (Figure 1). The minimum effective dose (MED) was defined as the minimal dose required to reduce the MI to 0.3.
Figur 1. Correlation between percentage of morphological change and metabolic activity index in CCL-1 cells. CCL-1 cells were treated with various doses of ASL and incubated for 48 h. After incubation, a number of rounding cells was counted, and then AlamarBlue was added to the cells. Metabolic activity index (MI) was calculated as described in Materials and methods. Pearson’s correlation of coefficient (r) was 0.83.
Immunization
Specific-pathogen-free six-week-old female mice were immunized with 10 µg of antigens with or without ASL by injection into the subcutaneous tissue of the inguinal region. At 3 weeks after immunization, Blood was drawn under anesthesia and the spleen was harvested after euthanasia. Bovine γ-globulin (Sigma-Aldrich, St Louis, MO, USA) and ovalbumin (OVA, Sigma-Aldrich) were used as the antigens.
Histopathological analysis
The mice were inoculated with 10 µL of ASL into the hind footpad. At 21 days after inoculation, mice were euthanized and footpad tissues were removed for histopathological examination. Samples of the hind footpads were fixed in neutral buffered formalin and decalcified in 5% EDTA (pH 7.4) at 4°C. The tissues were embedded in paraffin. Tissue sections (4-µm thickness) were stained with hematoxylin and eosin.
Quantitation of antibodies by enzyme-linked immunosorbent assay (ELISA)
Wells of a 96-well plate were coated with 100 µL of antigen solution at a concentration of 1 µg/mL in 50 mM sodium carbonate coating buffer (pH 9.6) for 18 h at 4°C. After three washes with Dulbecco’s modified phosphate buffered saline without magnesium and calcium (DPBS) contained 0.05% (v/v) Tween 20 (washing buffer), 300 µL of washing buffer that contained 10% (w/v) nonfat dried milk (incubation buffer) were added and the wells were incubated for 30 min at room temperature. After three washes with the washing buffer, two-fold serial dilutions of serum were added and incubated at room temperature for 2 h. After three washes, 100 µL of 5,000 times diluted Peroxidase-AffiniPure goat Anti-Mouse IgG (Jackson Laboratory, Bar Harbor, MA, USA) in the incubation buffer were added and incubated at room temperature for 2 h. The wells were then washed and 100 µL of substrate (TMB substrate kit; Thermo scientific, Waltham, MA, USA) were added. After incubation for 3 min in the dark at room temperature, 100 µL of 2 M H2SO4 were added to each well and the OD450 was measured. AP-AffiniPure goat Anti-mouse IgG, Fc γ Subclass 1 (Jackson Laboratory), and AP-Affinipure Goat Anti-Mouse IgG, Fc γ Subclass 2a Specific (Jackson Laboratory), were used to determine titers of IgG1 and IgG2a, respectively. For the assay with the alkaline phosphatase-conjugated secondary antibodies, FAST-pNPP kit (Sigma Aldrich) and 3 M NaOH (stopping solution) were used. In this case, OD405 was measured.
Antibody concentrations were expressed as the reciprocal of the serum dilution (titer) that gave an absorbance equal to the mean plus three standard deviations (SD) of the values from the negative control wells.
Statistical analysis
Statistical analysis was performed using the GraphPad Prism software version 6 (GraphPad Software, LA Jolla, CA, USA). The statistical difference between results from different groups was examined by ANOVA and Dunnett’s test.
Results
ASL suppresses the production of serum IgG against co-administered antigens
We examined serum IgG response of mice immunized with a mixture of bovine γ-globulin and ASL. Mice were injected with a mixture of 10 µg of bovine γ-globulin and 1,000 MED of ASL into the subcutaneous tissue of the right inguinal region. Sera from mice were collected 21 days after injection, and their bovine γ-globulin-specific serum IgG titers were measured by ELISA. The levels of bovine γ-globulin-specific IgG titer in sera from the mice injected with bovine γ-globulin alone were high (Figure 2). In contrast, the titers in sera from the mice injected with a mixture of the antigen and ASL were below the limit of detection (<50) (Figure 2), suggesting that ASL suppressed production of IgG against the co-administered antigen. On the other hand, when the antigen and ASL were injected into different sites, i.e., bovine γ-globulin and ASL were injected into right and left inguinal region, respectively, high titers of bovine γ-globulin-specific serum IgG were detected (Figure 2). The observation suggested that ASL suppressed antibody production only against co-administered antigen, but not against antigen administered in a different site from the site of ASL’s injection. When we repeated this experiment using OVA as an antigen, ASL suppressed serum antibody response against OVA (date not shown), suggesting that the immunosuppressive effect of ASL did not depend on the antigen.
Figur 2. Titers of γ-globulin-specific IgG in sera from mice administered with an antigen and ASL
Suppression of antibody response depends on mycolactone
We prepared ASL from a mycolactone-deficient strain ATCC19423 and determined its effect on serum IgG production to a co-administered antigen. Mice were immunized with 10 µg of bovine γ-globulin mixed with ASL extracted from M. ulcerans TMC1615 (284.7 µg of the ASL containing 1,000 MED) or ATCC19423 (284.7 µg of the ASL containing no detectable mycolactone). Serum IgG response to bovine γ-globulin was not detected in the sera from mice immunized with bovine γ-globulin mixed with the mycolactone-containing ASL prepared from strain TMC1615 (Figure 3). However, high titers of anti-bovine γ-globulin IgG were detected in the sera of mice immunized with bovine γ-globulin with ASL containing no mycolactone (Figure 3). The data suggested that the suppressive effect of ASL on antibody production depended on the presence of mycolactone.
Figur 3. The suppression of the antibody response against co-administered antigen is likely caused by mycolactone
Mycolactone-containing fraction does not induce the systemic suppression of antibody production
Mice were injected with 10 µg of bovine γ-globulin mixed with 1,000 MED of ASL into subcutaneous tissue of the right inguinal region, and 10 µg of OVA into that of the left inguinal region. Titers of bovine γ-globulin and OVA specific serum antibodies were measured three weeks after the immunization. The level of γ-globulin-specific IgG was below the limit of detection (Figure 4A). In contrast, the level of OVA-specific IgG was high, and there was no significant difference compared to the level of anti-OVA IgG in mice immunized with OVA only (Figure 4B). This result demonstrated that mycolactone-containing fraction did not affect antibody response against the antigen that was injected into a different site. The observation suggested that mycolactone did not affect systemic antibody production.
Figur 4. Localized suppression of antibody response by mycolactone-containing lipids fraction
The suppressive effect of mycolactone-containing fraction on antibody production is dose-dependent
We determined the titers of bovine γ-globulin-specific IgG in the sera from mice injected with 10 µg of bovine γ-globulin mixed with various doses of ASL (0, 10, 100, and 1,000 MED). As expected, a dose-dependent response was observed when the dose varied from 10 to 1,000 MED (Figure 5A). The titers of bovine γ-globulin-specific IgG in sera from mice immunized with 10 MED or more of ASL were significantly lower than that in sera from mice immunized with bovine γ-globulin only. The titer in the immunized group with 1,000 MED of ASL was below the limit of detection.
We also examined the level of bovine γ-globulin-specific IgG1 and IgG2a in the sera from mice immunized with the antigen and various doses of ASL. The bovine γ-globulin-specific IgG1 titers varied in a dose-dependent manner (Figure 5B), similar to whole IgG responses. However, the titers of bovine γ-globulin-specific IgG2a were not detected in all groups (date not shown).
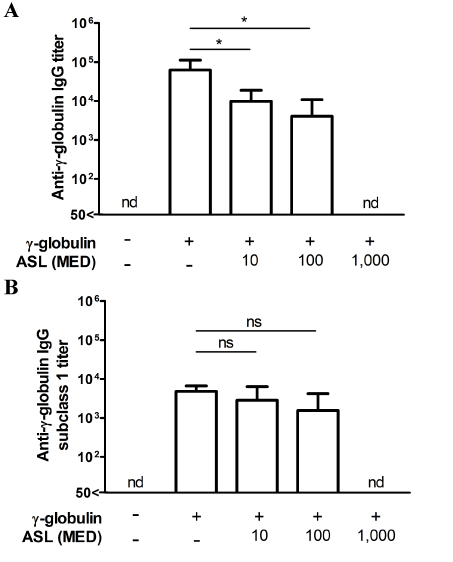
Figur 5. Dose-dependent suppression of antibody production by mycolactone-containing fraction
The immunosuppressive effect of mycolactone-containing fraction is observed below the toxic dose
Histopathology analysis was performed on mouse footpads injected with 10,000 MED of mycolactone-contaning fraction, which was ten-times higher than the dose that perfectly inhibited the antibody production against co-administered bovine γ-globulin. Mice were injected with or without 10,000 MED of ASL. Sections of the footpads were prepared as described in Materials and methods. No difference was observed in the histopathology of the footpad tissue between the two groups (Figure 6). In addition, ASL injection did not develop footpad swelling (data not shown).
Figur 6. Histopathological analysis of mouse footpads injected with ASL. Mice were injected with DPBS (A) and 10,000 MED of ASL (B). Three weeks after inoculation, histological sections of mouse footpads were stained with hematoxylin and eosin. Magnifications; ×100.
The observation suggested that mycolactone could affect antibody production under its toxic dose.
Discussion
Previous studies have reported that Th1 cytokine production in the serum of Buruli ulcer patients was suppressed compared to that in the healthy control group [17, 18]. In addition, several in vitro studies have suggested that mycolactone could inhibit cytokine and chemokine production from immune cells such as T cells, macrophages, and dendritic cells [9-13]. These observations clearly suggested that mycolactone presents immunosuppressive effects and may have an important role in the successful infection of M. ulcerans in humans. However, only few reports regarding the effect of mycolactone on serum immune responses are available in the literature so far. We previously reported that M. ulcerans-specific serum IgG was not detected in mice immunized with formalin-killed M. ulcerans cells [16]. However, when immunized with dewaxed whole-cells of M. ulcerans, mice produced high levels of M. ulcerans-specific serum IgG [16]. We hypothesized that mycolactone in the waxy cell wall of the bacteria may inhibit the production of M. ulcerans specific serum IgG.
In this study, ASL suppressed antibody production against co-administered bovine γ-globulin (Fig. 2) and OVA (data not shown). The effect was not observed if ASL from mycolactone-deficient M. ulcerans strain was used instead of ASL containing mycolactone (Fig. 3), suggesting that the effect was caused by mycolactone. The effect of mycolactone did not depend on antigens, and mycolactone only suppressed the antibody response against the co-administered antigen (Fig. 4). This observation suggested that the antibody suppression was based on the local activity of mycolactone at the site of injection and was not based on a systemic effect of mycolactone. Mycolactone might affect immune cells around the injection site and in the nearest peripheral lymph node (PLN). A previous study showed that large doses (50 or 100 µg) of mycolactone affected the crucial process of T cell homing in PLNs [19]. The inhibition of T cell homing by mycolactone might be one of the mechanisms underlying the suppression of serum antibody production. We measured IgG1 and IgG2a responses in mice injected with ASL and bovine γ-globulin in order to examine possible changes in the Th1/Th2 balance induced by mycolactone. However, IgG2a was not detected in any groups (Fig. 5). Phillips et al. suggested that mycolactone inhibited the production of IL-4 and IFN-γ from human T cells [11]. The inhibitory effect of mycolactone on the cytokine production could be linked to the suppression of antibody production, given that IL-4 is an important cytokine that induces Th2-type immune response. Interestingly, Ruf et al. showed the accumulation of B cells around infected sites of M. ulcerans in a mouse footpad model [20]. The accumulation of B cells could be caused by mycolactone, suggesting that mycolactone might also affect B cells directly or indirectly and disturb B cell activation and/or differentiation into antibody-producing plasma cells.
Recently, molecular targets of mycolactone were described. Wiskott-Aldrich syndrome protein (WASP) and neural WASP (N-WASP) were reported as direct binding targets of mycolactone [7, 21]. These proteins are members of a family of scaffold proteins transducing signals involved in the dynamic remodeling of the actin cytoskeleton [22-25]. Mycolactone exerts its effects by hijacking the WASP family, leading to uncontrolled activation of actin in the cytoplasm [21]. N-WASP is a negative regulator of B-cell signaling [23, 24], suggesting that the activation of N-WASP family by mycolactone could induce B-cell receptor (BCR) inactivation. Blocking of BCR signaling could be related to the suppression of antibody responses dependent on mycolactone.
On the other hand, other researchers reported that mycolactone inhibited the function of the Sec61 translocon [7, 9, 26]. Sec61 mediates antigen transport across endosomal membranes and affects cross-presentation in dendritic cells [27]. The inhibition of Sec61 by mycolactone could be one of the mechanisms underlying the suppression of antibody production observed in this study.
Angiotensin II type 2 receptors (AT2Rs) are also reported as mycolactone-binding proteins [7, 28]. AT2R blockers inhibit Th1 and Th2 immune responses in mice [29, 30]. Additionally, mycolactone affects RAW267.4, a macrophage-like mouse cell line, to cause hyperpolarization through AT2R signal inhibition [28]. This relationship between mycolactone and AT2R may also participate in the immunosuppressive role of mycolactone.
M. ulcerans exists in extracellular area, unlike other intracellular mycobacteria such as M. tuberculosis [16, 20]. Thus, specific antibodies to M. ulcerans might have a role in the clearance of M. ulcerans. M. ulcerans may use mycolactone to escape from immune responses in vivo by suppressing antibody production against itself.
Mycolactone-containing fraction suppressed the production of antibody only against the co-administered antigen, not against antigens administered into a different site, suggesting that mycolactone only has a local effect (Fig. 4). In addition, the dose that induced the antibody suppression showed no appreciable adverse effects based on histopathological analysis at least for 21 days following injection (Fig. 6). En et al. reported that injection with 100 µg of purify mycolactone induced significantly footpad swelling and loss of Schwann cell nuclei [31]. On the other hand, in our study, mouse footpads injected with 10,000 MED of ASL did not show any swelling (data not shown). The results suggested that En et al. might administer higher dose of mycolactone than our study. Further detailed evaluation of neurotoxicity of mycolactone might be necessary.
Taken together, mycolactone may be useful as a new immunoregulatory agent. Nano machines for medical use are being studied [32, 33]. Especially, protein-based nano machines are some of the most promising advances based on the accumulated knowledge on biologically active domains and sequences [32, 33]. Researchers could design artificial combination of such domains and easily prepare them by using any protein expression systems such as Escherichia coli. However, one critical problem of this strategy is the proteins’ antigenicity. Generally, immunoreactions could be developed in hosts against administered non-self-proteins, and the immune effectors, mainly antibodies, could inactivate (neutralize) the protein activity. Mycolactone could suppress antibody responses against co-administered protein-based nano machines, thereby improving the efficacy of protein-based therapy. Our results suggest that mycolactone might be useful when developing protein-based nano machines for medical use. For this purpose, safer derivatives of mycolactone will be necessary. We are currently studying the antigenicity of proteins conjugated with mycolactone. It would be interesting to investigate the mechanism for antibody suppression by mycolactone for possible use as some agent for future.
Conclusion
We demonstrated that partially purified mycolactone suppressed the production of specific antibodies against antigens co-administered with mycolactone. The suppressive effect of mycolactone-containing fraction was dose-dependent. Mycolactone was safe at a dose that was sufficient to induce the immunosuppression. Our results suggest that mycolactone may be useful as a new immunoregulatory agent.
Acknowledgments
We thank Naoya Tsukada, Sayuri Hishida, and Eri Suzuki-Kobayashi for skillful technical assistance. We are also grateful to Dr. Small (University of Tennessee, Knoxville, USA) for providing mycobacterial strains.
Author contributions
All authors contributed equally to this manuscript. NS designed the study, analyzed the data, and prepared the manuscript. HN assisted with data analysis and manuscript preparation. MW assisted with study design, data analysis, and manuscript preparation.
Funding informationThis study was supported by a grant from Research Organization for Infection Control Sciences of Kitasato University, Tokyo, Japan.
Competing interest
The authors declare that they have no competing interest.
References
- Sizaire V, Nackers F, Comte E, Portaels F (2006) Mycobacterium ulcerans infection: control, diagnosis, and treatment. Lancet Infect Dis 6: 288-296. [Crossref]
- Weir E (2002) Buruli ulcer: the third most common mycobacterial infection. CMAJ 166: 1691. [Crossref]
- Einarsdottir T, Huygen K (2011) Buruli ulcer. Hum Vaccin 7: 1198-1203. [Crossref]
- George KM, Chatterjee D, Gunawardana G, Welty D, Hayman J, et al. (1999) Mycolactone: a polyketide toxin from Mycobacterium ulcerans required for virulence. Science 283: 854-857. [Crossref]
- Kishi Y (2011) Chemistry of mycolactones, the causative toxins of Buruli ulcer. Proc Natl Acad Sci U S A 108: 6703-6708. [Crossref]
- George KM, Barker LP, Welty DM, Small PL (1998) Partial purification and characterization of biological effects of a lipid toxin produced by Mycobacterium ulcerans. Infect Immun 66: 587-593. [Crossref]
- Sarfo FS, Phillips R, Wansbrough-Jones M, Simmonds RE (2016) Recent advances: role of mycolactone in the pathogenesis and monitoring of Mycobacterium ulcerans infection/Buruli ulcer disease. Cell Microbiol 18: 17-29. [Crossref]
- Simmonds RE, Lali FV, Smallie T, Small PL, Foxwell BM (2009) Mycolactone inhibits monocyte cytokine production by a posttranscriptional mechanism. J Immunol 182: 2194-2202. [Crossref]
- Hall BS, Hill K, McKenna M, Ogbechi J, High S, et al. (2014) The pathogenic mechanism of the Mycobacterium ulcerans virulence factor, mycolactone, depends on blockade of protein translocation into the ER. PLoS Pathog 10: e1004061. [Crossref]
- Coutanceau E, Decalf J, Martino A, Babon A, Winter N, et al. (2007) Selective suppression of dendritic cell functions by Mycobacterium ulcerans toxin mycolactone. J Exp Med 204: 1395-1403. [Crossref]
- Phillips R, Sarfo FS, Guenin-Macé L, Decalf J, Wansbrough-Jones M, et al. (2009) Immunosuppressive signature of cutaneous Mycobacterium ulcerans infection in the peripheral blood of patients with buruli ulcer disease. J Infect Dis 200: 1675-1684. [Crossref]
- Boulkroun S, Guenin-Macé L, Thoulouze MI, Monot M, Merckx A, et al. (2010) Mycolactone suppresses T cell responsiveness by altering both early signaling and posttranslational events. J Immunol 184: 1436-1444. [Crossref]
- Pahlevan AA, Wright DJ, Andrews C, George KM, Small PL, et al. (1999) The inhibitory action of Mycobacterium ulcerans soluble factor on monocyte/T cell cytokine production and NF-kappa B function. J Immunol 163: 3928-3935. [Crossref]
- Adusumilli S, Mve-Obiang A, Sparer T, Meyers W, Hayman J, et al. (2005) Mycobacterium ulcerans toxic macrolide, mycolactone modulates the host immune response and cellular location of M. ulcerans in vitro and in vivo. Cell Microbiol 7: 1295-1304. [Crossref]
- Schütte D, Um-Boock A, Mensah-Quainoo E, Itin P, Schmid P, et al. (2007) Development of highly organized lymphoid structures in Buruli ulcer lesions after treatment with rifampicin and streptomycin. PLoS Negl Trop Dis 1: e2. [Crossref]
- Watanabe M, Nakamura H, Nabekura R, Shinoda N, Suzuki E, et al. (2015) Protective effect of a dewaxed whole-cell vaccine against Mycobacterium ulcerans infection in mice. Vaccine 33: 2232-2239. [Crossref]
- Zavattaro E, Mesturini R, Dossou A, Melensi M, Johnson RC, et al. (2010) Serum cytokine profile during Mycobacterium ulcerans infection (Buruli ulcer). Int J Dermatol 49: 1297-1302. [Crossref]
- Gooding TM, Johnson PD, Smith M, Kemp AS, Robins-Browne RM (2002) Cytokine profiles of patients infected with Mycobacterium ulcerans and unaffected household contacts. Infect Immun 70: 5562-5567.
- Guenin-Macé L1, Carrette F, Asperti-Boursin F, Le Bon A, Caleechurn L, et al. (2011) Mycolactone impairs T cell homing by suppressing microRNA control of L-selectin expression. Proc Natl Acad Sci U S A 108: 12833-12838. [Crossref]
- Ruf MT, Schütte D, Chauffour A, Jarlier V, Ji B, et al. (2012) Chemotherapy-associated changes of histopathological features of Mycobacterium ulcerans lesions in a Buruli ulcer mouse model. Antimicrob Agents Chemother 56: 687-696. [Crossref]
- Guenin-Macé L, Veyron-Churlet R, Thoulouze MI, Romet-Lemonne G, Hong H, et al. (2013) Mycolactone activation of Wiskott-Aldrich syndrome proteins underpins Buruli ulcer formation. J Clin Invest 123: 1501-1512. [Crossref]
- Silvin C, Belisle B, Abo A (2011) A role for Wiskott-Aldrich syndrome protein in T-cell receptor-mediated transcriptional activation independent of actin polymerization. J Biol Chem 276: 21450-21457.
- Becker-Herman S, Meyer-Bahlburg A, Schwartz MA, Jackson SW, Hudkins KL, et al. (2011) WASp-deficient B cells play a critical, cell-intrinsic role in triggering autoimmunity. J Exp Med 208: 2033-2042. [Crossref]
- Liu C, Bai X, Wu J, Sharma S, Upadhyaya A, et al. (2013) N-wasp is essential for the negative regulation of B cell receptor signaling. PLoS Biol 11: e1001704. [Crossref]
- Cotta-de-Almeida V, Westerberg L, Maillard MH, Onaldi D, Wachtel H, et al. (2007) Wiskott Aldrich syndrome protein (WASP) and N-WASP are critical for T cell development. Proc Natl Acad Sci U S A 104: 15424-15429. [Crossref]
- McKenna M, Simmonds RE, High S (2016) Mechanistic insights into the inhibition of Sec61-dependent co- and post-translational translocation by mycolactone. J Cell Sci 129: 1404-1415. [Crossref]
- Zehner M, Marschall AL, Bos E, Schloetel JG, Kreer C, et al. (2015) The translocon protein Sec61 mediates antigen transport from endosomes in the cytosol for cross-presentation to CD8(+) T cells. Immunity 42: 850-863. [Crossref]
- Marion E, Song OR, Christophe T, Babonneau J, Fenistein D, et al. (2014) Mycobacterial toxin induces analgesia in buruli ulcer by targeting the angiotensin pathways. Cell 157: 1565-1576. [Crossref]
- Chang Y, Wei W (2015) Angiotensin II in inflammation, immunity and rheumatoid arthritis. Clin Exp Immunol 179: 137-145. [Crossref]
- Mazzolai L, Duchosal MA, Korber M, Bouzourene K, Aubert JF, et al. (2004) Endogenous angiotensin II induces atherosclerotic plaque vulnerability and elicits a Th1 response in ApoE-/- mice. Hypertension 44: 277-282.
- En J, Goto M, Nakanaga K, Higashi M, Ishii N, et al. (2008) Mycolactone is responsible for the painlessness of Mycobacterium ulcerans infection (buruli ulcer) in a murine study. Infect Immun 76: 2002-2007. [Crossref]
- Dastjerdi R, Montazer M (2010) A review on the application of inorganic nano-structured materials in the modification of textiles: focus on anti-microbial properties. Colloids Surf B Biointerfaces 79: 5-18. [Crossref]
- Hamdi M, Ferreira A (2009) Multiscale Design and Modeling of Protein-based Nanomechanisms for Nanorobotics. Int J Robot Res 28: 436-449.