The article concerns application of embryonic model of Xenopus laevis for elaborating preparations directed either to suppression of cell proliferation, or, in opposite, to promoting forced cell differentiation. In the 1st series of studies the dynamics of the serotonin-modulating anticonsolidation protein (SMAP), being in linear relations with serotonin, in the embryos of Xenopus laevis throughout the stages of embryogenesis and metamorphosis was pursued with application of indirect ELISA-test. Beginning from the blastula stage till the end of the neurula its level remained unchanged. Thereafter continuous downregulation of the SMAP level, interrupted as a slight upregulation between 37th stage and onset of the 39th stage, was observed. In the 2nd series of studies incubation of the embryos of Xenopus laevis on the blastula and gastrula stages in fresh water containing SMAP at a dose of 50 and 100 μg/ml led to delay in development and, finally, to death of all the embryos within 4 days of observation. In the 3rd series of studies blocking of SMAP activity with the anti-SMAP polyclonal antibodies at a dose of 50 μg/ml in the embryos of Xenopus laevis, being on the 37th stage of development, resulted in their passing ahead (by two stages earlier) of the metamorphosis stage relatively to the rate of passing of this stage by the animals of the intact and control groups. So, if on the initial stages of embryogenesis SMAP realizes cytostatic activity, on the metamorphosis stage blocking its activity with antibodies, conversely, leads to passing ahead cell differentiation.
serotonin-modulating anticonsolidation protein (SMAP), antibodies, embryo¬genesis, metamorphosis, Xenopus laevis
Certain stages of embryonic processes have high level of resemblance with carcinogenesis processes, particularly in high proliferative activity of their composing cellular elements. Along with it, the advantage of studying embryonic stages relatively to cancer processes concludes in that that processes of cell proliferation and differentiation are diverged over time, mostly within different embryonic stages. So, utilization of embryonic models makes possible to elaborating the approaches directed purposefully and separately on the processes of cell proliferation and differentiation. Issuing from high proliferative potential of the cancer cells, it is clear why suppressive effects on cell proliferation and the remedies possessing with such activities are under meticulous attention of the most researches dealing with the studies of carcinogenesis and seeking ways of combating with tumor. Along with it, from the first glance, the unusual interest to the processes of cell differentiation and study of the underlying molecular mechanisms, coming from the cancer-absorbed researchers, is related to a problem of the cancer stem cells constituting serious obstacle in treatment of malignant tumors [1-3]. The problem of their eradication is complicated by the fact that broadly applied chemotherapeutic preparations are ineffective against these cells. Sometimes the struggle with cancer stem cells is likened to the struggle with weeds: you can chop off the weeds’ stalks many times, but they will grow again and again, so far, their roots still remain in the soil. The same is referred to the cancer stem cells in the sense that the role of woods’ stalk in this case belongs to mature malignant tumor cells, while the role of woods’ roots – to the cancer stem cells themselves.
The goal of the present study was application of different stages of embryonic and early stages of development of Xenopus laevis for the purpose of elaborating preparations directed either to suppression of cell proliferation, or, in opposite, to promoting forced cell differentiation. This goal was achieved through application of serotonin-modulating anticonsolidation protein (SMAP) and anti-SMAP polyclonal antibodies for the purpose of blocking its activity in different stages of the embryonic and early stages of development of Xenopus laevis.
Biochemical techniques
SMAP, being in linear relations with serotonin, was purified from the cow brains with application of two-step purification procedure as had been described earlier [4]: 1) partial precipitation with sodium sulfate in the range of 0-40% concentration; 2) gel-chromatography on the column (3.0 X 60.0 cm) of Sephadex G-150. SMAP purification was carried out under the screening control of the indirect ELISA-test [5] with application of anti-SMAP rabbit immunoglobulins to selecting the SMAP-enriched protein fractions. The protein purity was checked by electrophoresis in polyacrylamide gel.
The anti-SMAP polyclonal immunoglobulins were produced through immunization of the rabbits with SMAP, using 300 μg of the protein always in mixture with the complete Freund adjuvant. SMAP and anti-SMAP immunoglobulins were frozen and kept under -70○C.
The content of SMAP was determined in the embryos and tadpoles of Xenopus laevis by the indirect ELISA-test realized on the polystyrene plates of moderate adsorption [5]. The samples were homogenized and water-soluble proteins were extracted and used as antigens in the ELISA-test. Specific polyclonal rabbit anti-SMAP antibodies were used as the first antibodies, while the anti-rabbit goat immunoglobulins, coupled with covalent bonds with horseradish peroxidase, were used as the second antibodies. Orthophenylendiamine was used as a substrate for peroxidase to visualize the results of the reaction. The reaction was stopped by adding of 3 M NaOH and the results were transformed into digital form by the ELISA-test reader of the model of “Molecular Devices Spectra Max 250” (MTX Lab Systems, Inc., USA) at the wavelength of 492 nm (wavelength of reference 630 nm) and analyzed with application of the t-Student criterion.
Embryonic technique
The embryonic studies were conducted on Xenopus laevis. The eggs from the sexually mature animals were obtained after injecting them with human gonadotropin at a dose of 150 units for males and 350 units for females.
In the 1st series of studies after several hours since gonadotropin administration, females released roe which was fertilized by males. Simultaneously after roe fertilization samples of the embryos were taken from different stages of embryonic development (stages 1-2, 7, 9, 11, 13, 20, 22, 28), pre-metamorphosis, metamorphosis and post-metamorphosis (stages 37, 42, 44, 45, 49, 53) for evaluation of the level of SMAP with application of the indirect ELISA-test and anti-SMAP polyclonal immunoglobulins. For evaluation of SMAP 10 samples of specimens within each stage were taken. The results were averaged within each group and differences between adjacent stages were evaluated on t-Student’s criterion.
In the 2nd series of studies the embryos, being on the blastula and gastrula stages, were placed into the Petri dish with fresh water containing different preparations. The embryos were culled into three groups: 1) intact group (n = 17), 2) control group – SMAP and anti-SMAP antibodies – both at a concentration of 50 μg/ml (n = 15), and 3) experimental group – SMAP at a concentration of 50 and 100 μg/ml (n = 17).
In the 3rd series of studies the animals, being on the 37th stage of development (beginning from the late tailbud stage), were placed into the Petri dish with fresh water containing different preparations. The embryos were culled into four groups: 1) intact group (n =13), 2) control group – SMAP and anti-SMAP antibodies - both at a concentration of 50 μg/ml (n = 13), 3) 1st experimental group – SMAP at a concentration of 50 μg/ml (n = 13), 4) 2nd experimental group – SMAP at a concentration of 100 μg/ml (n = 13), and 5) 3rd experimental group – anti-SMAP antibodies – at a concentration of 50 μg/ml (n = 13). After 24 h the animals were transferred into the tanks with fresh water with regulated temperature and air supply. On the basis of elaborated tables of early stages of development of Xenopus laevis [6] registration of passing metamorphosis stage by the animals of different groups was carried out. Particularly, definition of passing of this stage by the animals was realized on the basis of degree of the tail resorption, advent of the precursors (buds) of the hind legs, changes in the shape of mouth and other second-order morphological changes typical to mature animals.
The results were averaged within each group and differences between different groups were evaluated on Wilkoxon-Mann-Whitney’s U-criterion.
Dynamics of SMAP level on embryonic stages and metamorphosis in Xenopus laevis
In the 1st series of studies the results revealed the dynamics of the level of SMAP in the organism of the animals throughout the early stages of ontogenesis (embryogenesis and metamorphosis) of Xenopus laevis. It was noticed that after slight downregulation of SMAP immediately after fertilization of the roe, on the forthcoming stages till finalization of the neurula its level remained unchanged, on the same values. Thereafter continuous and significant (p < 0.01 and p < 0.001) downregulation of the SMAP level, interrupted once in the form of its slight upregulation in the interval between middle of the late tailbud stage (37th stage) and onset of the pre-metamorphosis stage (39th stage; Figure 1), was observed.
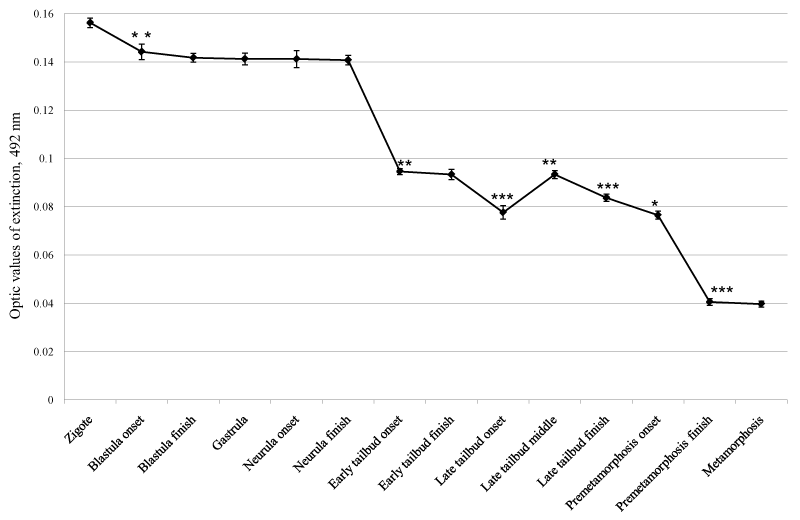
Figure 1. Changes of SMAP level in early stages of ontogenesis in Xenopus laevis. **- p < 0.01; *** - p < 0.001 relatively to preceding point.
Role of SMAP in regulation of embryonic development and metamorphosis in Xenopus laevis
In the series of studies incubation of the embryos of Xenopus laevis on the blastula and gastrula stages in fresh water containing SMAP at a dose of 50 and 100 μg/ml led to delay of their development and, finally, to death of all the embryos of this group within 4 days of observation. At the same time the embryos of the intact and control groups left alive and developed normally, without lethality of any specimens.
In the of studies blocking of SMAP activity with the anti-SMAP polyclonal antibodies in the experimental group realized significant effect on the rate of animal passing through embryogenesis. Particularly, it was shown that single addition of the anti-SMAP antibodies at a dose of into the incubation milieu of the embryos of Xenopus laevis, being on the of development, after 14 days resulted in their passing ahead (two stages earlier) of the metamorphosis stage (50th stage) relatively to the rate of passing of this stage by the animals of the intact (and control groups (p = 0.01). Along with it, animals of the and experimental groups under effects of SMAP of both doses were on the 49th stage of development passing ahead metamorphosis by one stage earlier relatively to the animals of intact and control groups (p = 0.01).
So, up to the end of the neurula stage of embryogenesis of Xenopus laevis the level of SMAP in the organism remains unchanged, while its continuous downregulation on the next stages is observed. Addition of SMAP to the embryos at the stages of blastula and gastrula leads to delay of embryonic development and, finally, to death of all the embryos. Blocking of SMAP activity by anti-SMAP antibodies on the stage of pre-metamorphosis of Xenopus laevis leads to noticeable passing ahead of the metamorphosis stage by the embryos.
The results of the studies indicate to close involvement of SMAP, realizing serotonin functions on sub-cellular level, in the processes of cell proliferation and differentiation. As the initial stages of embryogenesis are characterized with high indexes of cell proliferation, remaining of the SMAP level stable, in the form of smooth horizontal line during theses stages up to the end of neurula stage shows that such unchanged amount of SMAP is required for molecular support of intensive cell proliferation.
As it is known from the literature, on the next after neurula stages of embryogenesis and initial stages of development including metamorphosis, decline of cell proliferation and, in opposite, step-by-step strengthening of cell differentiation are observed [6]. Gradual downregulation of SMAP on the said stages indicates that its level is synchronized negatively to realization and/or regulation of differentiation processes, thus giving grounds to making a conclusion that SMAP itself is engaged in negative regulation of cell differentiation, probably, through switching on differentiation-launching genes on the background of SMAP downregulation.
The above stated conclusions, issuing from the results of the of the studies, were confirmed later by the results of the undertaken . Particularly, artificial upregulation of the SMAP level in the embryos through its addition to the incubation milieu of embryos, staying on the blastula and gastrula stages, under the both applied doses brought to cessation of the embryo development and total death of the embryos. So, SMAP upregulation on the initial stages of embryogenesis realizes cytotoxic effects on the embryonic cells.
The observed cytotoxic effects of SMAP on the onset of embryogenesis of Xenopus laevis may be related to induction by SMAP of conformational changes of chromatin transducting it into the condensed inactive form. This idea is confirmed by our ealier studies in which SMAP administration to the sturgeon juveniles led to significant downregulation (by over 50%) of the level of mutagenic changes in the somatic cells induced by soil sediments from Baku Bay containing high levels of heavy metals and polyaromatic hydrocarbons relatively to the control group kept under the similar polluted conditions. In the control animals this contamination induced 5-times elevation of mutations relatively to the intact animals kept in fresh water [7]. These earlier obtained data give grounds to make a conclusion that SMAP brings chromatin to the condensed, folded state, this way providing its protection from the effects of adverse factors.
Along with it, antibodies-mediated blocking of SMAP activity on the 37th stage of development significantly fomented passing ahead metamorphosis (by two stages earlier) by the embryos of the 3rd experimental group relatively to the intact and control animals, this way supporting the above proposed idea of existing significant negative regulation of the differentiation processes of embryonic cells by SMAP, issuing from its downregulation on post-neurula stages of development. Besides, such passing ahead of metamorphosis, though in somehow fainter degree (by one stage earlier), was noticed for the effects of SMAP at both studied doses. However, these effects of SMAP itself are, probably, due to the phenomenon of down-regulation of SMAP-accepting receptors resulting from their internalization into the embryonic cells under the effect of applied doses of SMAP. So, these results give grounds in the future for application of anti-SMAP antibodies for the purpose of forced differentiation of immature embryonic-resembling cells, particularly cancer stem cells, having many common features with the embryonic cells. The idea of possible impact on cancer stem cells through modulating microenvironmental stimuli, leading to their differentiation, has been put forward by different researchers [3,8,9].
- Fulawka1 L, Donizy P, Halon A (2014) Cancer stem cells – the current status of an old concept: literature review and clinical approaches. Biological Research 47: 66-75.
- Pan Q, Li Q, Liu S, Ning N (2015) Concise Review: Targeting Cancer Stem Cells Using Immunologic Approaches. Stem Cells 33: 2085-2092. [Crossref]
2021 Copyright OAT. All rights reserv
- Cojoc M, Mäbert K, Muders MH, Dubrovska A (2015) A role for cancer stem cells in therapy resistance: cellular and molecular mechanisms. Semin Cancer Biol 31: 16-27. [Crossref]
- Mekhtiev AA (2000) Revealing in the rat brain protein, possessing with anticonsolidation properties. Bulletin of Exper Biol Med 129:147-150.
- Catty D (1991) Antibodies. A Practical Approach, Oxford University Press, Oxford, New York, Tokyo, p. 384.
- Sive HL, Grainger RM, Harland RM (2010) Early Development of Xenopus laevis. A Laboratory Manual Cold Spring Harbor Laboratory Press. Cold Spring Harbor, New York p. 340.
- Mekhtiev AA, Movsum-zadeh SK (2008) Antimutagenic activity of serotonergic system and underlying mechanisms in sturgeon juveniles (Acipenser guelden¬staedti persicus) and goldfish (Carassius auratus). J Evol Biochem Physiol 44: 476-481.
- Radosevich J, Aqil M, Bassiony M (2015) Rethinking the tumor stem cell theory. Tumor Biology 36: Suppl. 1: S24.
- Sell S (2004) Stem cell origin of cancer and differentiation therapy. Crit Rev Oncol Hematol 51: 1-28. [Crossref]