Recent investigations [1] of EPR (Einstein-Podolsky-Rosen)-generated [2] entangled proton qubit-pairs demonstrate that Grover’s [3] enzyme quantum-readers quantify quantum informational content embodied within entangled proton qubit genomic systems [4]. Double-helical DNA is replicated into a “metastable” keto-amino state; consequently, metastable hydrogen bonded amino (–NH2) genome protons encounter quantum uncertainty limits [1,4], Δx Δpx ≥ ħ/2. The resulting quantum mechanical proton – proton interaction introduces probabilities of EPR-arrangements, keto-amino ―(entanglement) → enol-imine, yielding “qubit-pairs” of entangled enol and imine protons (Figures. 1,2). Reactions yielding EPR-generated entangled proton qubit-pairs are observed [1,4] as G-C → G´-C´, G-C → *G-*C and A-T → *A-*T. (Bold italics ― G´-C´, *G-*C, *A-*T ― denote necessity of Hilbert space to describe dynamics of embedded entangled proton qubit-pairs; see Figures. 1&2 for notation.)
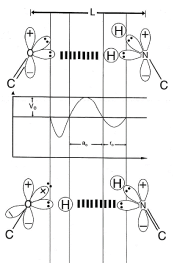
Figure 1. Schematic of “metastable” keto-amino and “ground state” enol-imine hydrogen bonds
Amino protons encounter quantum uncertainty limits, ΔxΔpx ≥ ħ/2, introducing probabilities of EPR arrangements, keto-amino ― (entanglement)→ enol−imine. The asymmetric double-well potential represents an energy surface “seen by” metastable hydrogen bonding amino proton, and a “ground state”, entangled enol and imine proton “qubit-pair”. Product enol and imine protons are entangled, and are each shared between two indistinguishable sets of electron lone-pairs, and thus, participate in entangled quantum oscillations, │+> ⇄ │─>, between near symmetric energy wells, occupying intramolecular decoherence-free subspaces.)
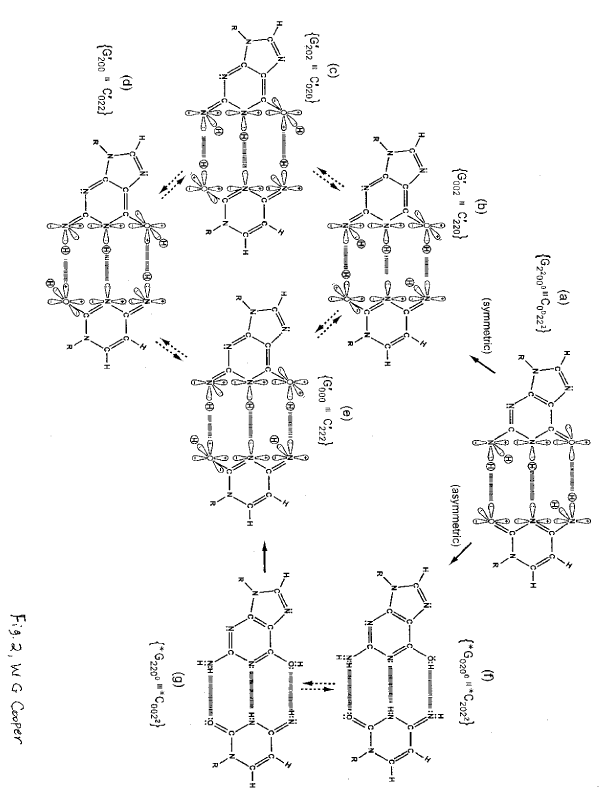
Figure 2. Distribution of entangled proton qubit states at a G′-C′ (symmetric) or *G-*C (asymmetric) superposition site.
Dynamic, “intra-atomic”, EPR-generated entangled proton qubit states, │+> ⇄ │–>, occupy decoherence-free subspaces [1,4], and thus, are resistant to decoherence by “normal” in vivo classical environments [1,4]. Grover’s [3] enzyme quantum reader “measures”, δt << 10–13 s, quantum informational content by “trapping” a qubit state,│–>, within a genome groove [4]. These measurements, ẟt << 10‒13 s, provide instructions for executing quantum information processing, Δt́ ≤ 10‒14 s [4], of entangled proton qubits, which specifies evolutionary distributions of the 22 most abundant rat and human microsatellites [1,5]. Since the evolutionary “emergence-distance” between rat and human is ~ 70´106 y [6], EPR-generated entangled proton qubit-pairs must retain entangled “two-state” quantum coherence, │+> ⇄ │–>, for years to decades, before specifying microsatellite evolution instructions with “measured” quantum informational content of EPR-generated entangled proton qubit-pairs [1,4]. Otherwise, Grover’s-type [3] quantum processor measurements, δt << 10–13 s, of EPR-generated entangled proton qubit-pairs could not accurately specify evolutionary distributions of the 22 most abundant microsatellites common to rat and human [5], as exhibited [1,4,5].
These results [1,4-6] imply that EPR-generated entangled proton “qubit-pairs” [2] are resources for quantum information processing in all double helical DNA systems, including human brain-cell DNA [1,7]. In this case, quantum information processing, Δtʹ ≤ 10–14 s [1,3,4], of EPR-generated entangled proton qubits, executed by a single brain-cell, could communicate the resulting quantum information processing calculations to the brain’s neuronal network of ~ billons of neurons [7]. This evolutionarily coordinated “network” communication could cause coupled, entanglement-enabled information-processing computations in ~ billons of brain-cell DNA systems, and consequently, could execute the phenomenon of consciousness [7,8]. A model for Homo sapiens’ consciousness [1,7] is thus implied by ~ 3.6´109 y of entanglement-enabled evolution [4], from duplex RNA–ribozyme segments, into double helical DNA of ~ 6.8´109 bp, which resides in Homo sapiens’ brain-cells [1,4].
Homo sapiens’ brain contains sensory “biological files” [1,7] for vision, acoustics, vocal, olfactory, touch, taste, balance, self-motion, pain, emotion, language, analyses, music, imagination, “truth” and “fantasy”. Sensory files are interfaced with, initially vacant, “hard drive” memory files that acquire input data from one or all sensory files. Brain-cell DNA of robust human infants has been evolutionarily selected to accumulate “stable” EPR-generated entangled proton qubits [1], for purposes of storing quantum information, and executing its quantum processing [4]. Brain-cell “DNA reservoirs” are incrementally populated with EPR-generated entangled proton qubits to their “threshold limits” [1,4], which is achieved by age ~25 y for H. sapiens. Consciousness is a consequence of Grover’s processors continually conveying quantum informational instructions for consciousness-relevant quantum information processing of entangled proton qubits. Anesthetics [8] inhibit Grover’s processors, which cause rapid transitions from consciousness to unconsciousness. This explanation implies “normal”, non-drug-induced sleep is a consequence of “inactive” Grover’s [3] processors, which ‒ during consciousness ‒ actively measure quantum informational content of EPR-generated entangled proton qubits. Thus, normal sleep is a “down-time” condition that allows EPR-generated entangled proton qubits to repopulate “recently measured” segments of double helical DNA [1,4], while Grover’s processors are “inactive”. In this case, the “sleeping”, unconsciousness brain is operating on “autopilot”, without Grover’s processors continuously “reading” quantum informational content embodied within EPR-generated entangled proton qubits, which provides instructions for consciousness-relevant quantum information processing.
- Cooper WG (2017) EPR–proton qubits’ role in evolution and age-related disease. Phys Sci & Biophys J 1: 000104. [Crossref]
- Einstein A, Podolsky B, Rosen N (1935) Can Quantum Mechanical Description of Physical Reality be Considered Complete? Phy. Rev 47: 777–780. [Crossref]
- Grover LK (1997) A fast quantum mechanical algorithm for database search. Phys Rev Lett 79: 325. [Crossref]
- Cooper WG (2018) Consequences of EPR-proton qubits in DNA. Adv Quantum Chem 77.
- Beckmann JW, Weber JL (1992) Survey of human and rat microsatellites. Genomics 12: 627–631. [Crossref]
- Kumar S, Hedges SB (1998) A molecular timescale for vertebrate evolution. Nature 392: 917-920. [Crossref]
- Dehaene S, Lau H, Kouider S (2017) What is consciousness, and could machines have it? Science 358: 486-492. [Crossref]
- Hameroff S, Penrose R (2014) Consciousness in the universe: a review of the “Orch OR” theory. Physics of Life Reviews 11: 39-78. [Crossref]