Abstract
The article being a pioneer in medical abzymology, integrating basic innovations, biodesign-driven translational applications and clinical practice, summarizes and reviews the current knowledge and practice in the field, covering everything from theoretical aspects to practical considerations and including applications from organic synthesis to medical uses, uses the resources of bioinformatics. One of the most attractive and promising areas of medical abzymology are anti-body-proteases (or Ab-proteases combining exquisite specificity with permanent and po-tent antigen inactivation.
Ab-proteases belong to a new group of physiologically active substances with dual characteristics: they represent a pool of canonical autoAbs, endowing with targeted pro-teolytic activity, which is crucially important in the progression of clinical autoim-munity syndrome and inflammatory disorders. Ab-proteases are crucial for immune homeostasis regulation and can be of practi-cal value in the development of modern immunodiagnostic tools and schedules of targeted immunotherapy. The innovations and achievements in the latter supported by IT modelling, have enriched our deep understanding of life-related catalysis, particularly by bio-molecules in human bio fluids and liquid systems as a whole. The article is the first source to provide such comprehensive coverage and constitutes a prime reference for bio-researchers, lab technicians, bio designers, bioengineers, and precision pathologists, immunologists, and practitioners.
Keywords
abzymes, antibody-proteases, medical abzymology, biocatalysis, anri-body-mediated proteolysis, bioinformatics, IT modeling
Introduction
Catalytic antibodies (abzymes) have attracted attention because they can combine exquisite specificity with potent antigen inactivation, i.e., are featured with dual func-tionality. In reality, abzymes belong to a new group of physiologically active substances: they represent a pool of canonical autoantibodies (autoAbs), endowing with catalytic activity (Figure 1). In common with enzymes, Abs can specifically bind a large range of chemical structures and using the same type of molecular interactions, but, in contrast, the immunoglobulins (Igs) bind tightly to the target molecule in their ground state and then are unable to catalyze reactions. Abzymes (catalytic Abs) are part of the Ab’s repertoire in vivo.
Figure 1. Abzymes as antibodies (Abs) endowed with enzymatic activities
The concept that Abs may catalyze chemical reactions was pioneered by L. Pauling in the 40's, who suggested that an enzyme lowers the energy barriers of a reaction by stabilizing preferentially the transition state of the substrate during the reaction rather than the substrate in its ground state [1].Twenty years later, W. Jencks proposed a strategy to prepare new biocatalysts, which was based on the production of Abs directed against stable molecules resembling the transition state structure of a specific chemical transformation [2].
In addition to providing a strategy for generating catalysts for reactions difficult to achieve by existing enzymatic or chemical methods, Ab catalysis provides a tool to gain increased insight into the mechanisms of biocatalysis and the evolution of catalytic function [3-5].The years passed, gifted us numerous and unique achievements in a new area of biocatalysis, i.e. abzymology, being based on abzymes. This trend opens prospects for producing new types of biocatalysts with manifold properties for design-driven biotech and Personalized & Precision Medicine (PPM)-guided clinical practice [6]. The success that was born in experiments on the design of artificial abzymes, stimulated further the emergence of a stage of studying the physiological role of natural abzymes in the patho-genesis of major diseases, i.e., the emergence of the era of medical abzymology [7,8]. Whose achievements can be applied in the management and treatment of autoimmune and cancer conditions with pharmaceutic constructions designed on abzyme analogue base or abzyme-encoding gene in interventional immune or gene-driven immune therapy [9,10].
Natural proteolytic antibodies (antibody-proteases) as applicable in clinical practice
The discovery of natural catalytic Abs and generation of engineered abzymes has become a valuable step in developing abzymes to diagnose, to monitor, to control, to treat and to rehabilitate patients at clinical stages and to prevent the disorder at subclinical ones in persons at risks to secure the efficacy of regenerative manipulations. This information can allow to design the algorithms for combinatorial (preventive, prophylactic, therapeutic and rehabilitative) treatment, whilst developing unique tools for individually therapy for a number of diseases [11,12]. Among the abzymes discovered and studied, proteolytic Abs (Ab-proteases) are of special value. The latter are an important pathogenic factor in the progression of clinical autoimmunity syndrome, inflammatory disorders or infectious diseases [13-16,17-33].
Ab-proteases to monitor infectious diseases naturally occurring Ab proteases to microbial determinants have been correlated with resistance to infection and provoked substantial interest in harnessing the power of Ab-mediated proteolysis against microbial antigens for host defense. For instance, septicemia is the most common cause of death in intensive care units, and it is caused by the toxic host’s systemic response to infection. Abs have been proposed to be involved in the removal of metabolic waste and the prevention of infection the high levels of abzymes are associated with a good prognosis of septicemia [34]. Ab-proteases can reduce infection and post infection inflammation in septicemia patients but that a lack of catalytic Ab response may accelerate the occurrence of the dis-ease. It is a better potential way to treat septicemia. In this sense, the HIV-1 coat proteins gp120 and gp41 have been targeted because of the key role they play in infecting CD4 + T lymphocytes – those three Ab proteases are able to cleave gp120 in a site-specific manner were generated upon immunization of mice with a CRA-coupled gp120 [35].
Regarding COVID-19-associated abzymes, Y. Song et al have proposed that some patients with COVID-19 may produce Abs against SARS-CoV-2 with proteolytic activity that target important proteolytic regulatory cascades [36]. The discovery of “abzymes” and their translational applications could pave the way for new treatments targeting the acute effects of COVID-19, long COVID, and possibly other viruses. Ab proteases could become promising molecular tools not only in diagnostics, but also in treatment of a number of infectious diseases, participating in the formation of passive immunity due to its own functional resource - the ability to site-directed hydrolysis of target objects with the manifestation of cytotoxic properties.
Ab-proteases to monitor inflammatory and autoimmune diseases of tremendous value in this sense are Ab-proteases directly affecting the physiologic remodeling of tis-sues with multilevel architectonics (for instance, myelin, cardiac muscle or cancer tissues), whilst securing the requests and standards of regeneration. They can also cleave specific surface proteins on viruses or tumor cells, thereby disrupting the invaders. Rationales for the requirements of proteolytic Abs as therapeutics are still accumulated [18-33]. Proteolytic Abs isolated from a variety of clinical conditions are able to specifically hydrolyze thyroglobulin (TG), cardiac myosin (CM), myelin-basic protein (MBP), and factor VIII (FVIII) or factor IX (FIX), respectively. The therapeutic relevance of these findings is discussed [13]. In the overwhelming number of autoimmune thyroiditis (AIT) cases, such Ab proteases are concentrated among patients with hypothyroidism and a tendency to hypo- and, subsequently, to thyroid atrophy, and in such patients, as a rule, both types of Ab proteases are present (as part of serologic tandems). In this regard the screening procedure for TG-targeted Ab-proteases may be considered as a highly sensitive and specific indicator test in the diagnosis and prediction of AIT, which allows one to diagnose not only within the framework of major thyroid entities, but much broader, by covering a great variety of syndrome-related forms of clinical pathology.
The presence of anti-CM autoAbs in the blood of AIM patients is counteracted by their high proteolytic activity against CM as a specific substrate. CM-targeted Ab proteases show a direct correlation with both the severity of the clinical picture of AIM and the dosage of therapeutic drugs administered to the patient. The process of permanent destruction of cardiac muscle with high frequency of exacerbations is accompanied by bursts of Ab-protease activity. The latter may be one of the reasons for the formation of focal and diffuse lesions with the development of dilated cardiomyopathy (DCMP) and persistent heart failure, and thus serve as a significant pathogenic factor in the progression of autoimmune syndrome in myocarditis [16,37].
CM-targeted Ab proteases may also have a functional role in AIM-related preillness persons-at-risk, as suggested by in vitro data as well as by preliminary clinical observations, though further work is in progress to clarify this important issue. In this sense, of great interest is the evolution of Ab-associated proteolytic activity at different stages of AIM progression - the activity of Ab-proteases is first registered at the subclinical stages (4-12 months prior to the clinical illness), revealing significant correlation with scales of auto aggression and the disability of the patients with AIM. Therefore, the activity of Ab-proteases and its dynamics tested would confirm a high subclinical and predictive value of the tools as applicable for monitoring AIM at subclinical and clinical stages [38]. The core of our working hypothesis is that the degradation of cardiac muscle tis-sue under the action of AT-mediated proteolysis, which is essential for the progression of heart failure (HF), is the cause of persistent HF during the resolution of active inflammation and transformation of AIM into DCMP [39].
Bronchial asthma (BA) produces autoAbs that are also capable of Ab-mediated targeted proteolysis - they specifically bind vasoactive intestinal peptide (VIP) and hydrolyze it efficiently [40] (Figure 2). Proteolytic anti-MBP Abs were isolated from MS patients [18]. The activity of those MBP-targeted Ab-proteases markedly differs from healthy controls and correlates with the expanded disability status [41-42]. Some of the MBP-targeted Ab-proteases were found to demonstrate their highest activity in MS patients with the most severe (exacerbation or progression phases) or moderate (remission or stabilization phases) clinical courses (Figure 3A) or dependence of the incidence of MBP-targeted Ab-proteases on the EDSS score in MS patients (Figure 3B).
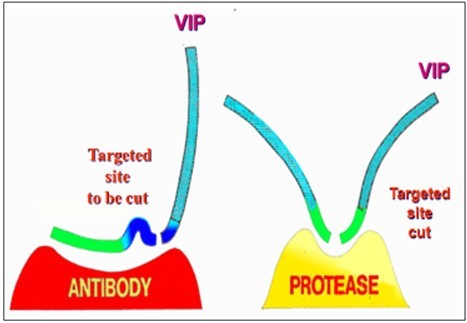
Figure 2. The anti-VIP proteolytic antibody binds a seven-amino acid subsequence of VIP distant from the cutting site (shown as a gap). Adapted from: 41
Figure 3A. The correlation of MBP-targeted Ab-associated proteolytic activity with scales of demyelination and neurological deficiency
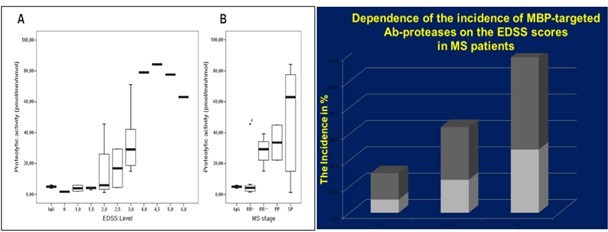
Figure 3B. Dependence of the incidence of MBP-targeted Ab-proteases on the EDSS score and thus with the disability of MS patients
The MBP-targeted Ab-associated proteolytic activity demonstrated also significant correlation with scales of demyelination, neurological deficiency and thus with the disability of the patients (it is seen from the EDSS scores) EDSS, The Expanded Disability Status Scale (EDSS) is a method of quantifying disability in multiple sclerosis and monitoring changes in the level of disability over time. It is widely used in clinical trials and in the assessment of people with MS. The EDSS scale ranges from 0 to 10 in 0.5 unit increments that represent higher levels of disability. EDSS (the Expanded Disability Status Scale (EDSS) is a method of quantifying disability in multiple sclerosis and monitoring changes in the level of disability over time. The MBP-targeted Ab-associated proteolytic activity demonstrated significant correlation with scales of demyelination, neurological deficiency and thus with the disability of the patients (it is seen from the EDSS scores).
Meanwhile, sequence specificity of MBP-targeted Ab proteases is the capacity to distinguish between particular epitopes in the MBP molecule. In this context, FIVE preferential sites of cleavage of the MBP molecule by abzymes isolated from MS patients were identified [43] (Figure 4). Ab-mediated proteolysis of MBP results in generating a set of peptides with MW ranged in various but fixed boundaries to suit common principles of the molecular architectonics of MBP. The final statistical data revealed FIVE sites of preferential proteolysis MS, multiple sclerosis; MB, myelin basic protein; Ab, antibody.
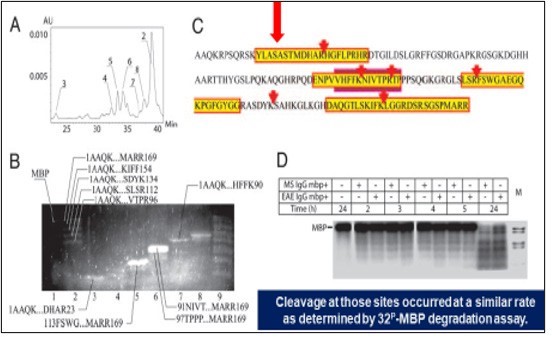
Figure 4. Sequence specificity of the MBP-targeted Ab-mediated proteolysis by anti-MBP au-toAbs harvested from MS patients
The progression of MS and/or in the aggressive forms, Ab-proteases would attack presumably highly immunogenic epitopes. The highly active Ab-proteases are typical for the stages with full-scale clinical manifestations or/and for MS aggressive types. The moderate activity is typical for less aggressive phases of MS, including subclinical stages. Registration in the evolution of highly immunogenic Ab-proteases would illustrate either risks of transformation of subclinical stages into clinical ones. Therefore, the proposed predictive value of MBP-targeted Ab- proteases for the development of MS is being challenged! And the activity of Ab-proteases and its dynamics tested would confirm a high subclinical and predictive value of the tools as applicable for PPM-guided monitoring protocols. Of special interest, is a protease-activated Ab (pro-Ab) targeting vascular cell adhesion molecule 1 (VCAM-1), a marker of atherosclerotic plaques, was constructed by tethering a binding site-masking peptide to the Ab via a matrix metalloprotease (MMP) susceptible linker. Pro-Ab activation in vitro by MMP-1 yielded a 200-fold increase in binding affinity and restored anti-VCAM-1 binding in tissue sections from ApoE (–/–) mice ex vivo. These results demonstrate that the activity of disease-associated Ab proteases can be exploited to site-specifically target Ab activity in vivo [44]. The phenomenon of targeted Ab-mediated catalysis is extremely interesting and can potentially be applied to many different objectives including new types of efficient catalysts, evaluation of the functional roles of abzymes in innate and adaptive immunity, and understanding of certain aspects of self-tolerance and of the destructive responses in autoimmune, inflammatory and infectious diseases.
Ab-proteases through the view of design-driven translational applications-Future perspectives
Ab-proteases and abzymes, globally, can continue to play a role and even fully prevent the emergence of autoimmune disorders, especially in the field of infection and immunity, where the process of its occurrence and development often takes a long time [4,5,45]. The development of artificial synthetic abzymes is still in the initial stage, and significant progress has been made after several years of efforts. Meanwhile, the drastic improvements in design-inspired and IT-supported bioengineering technologies, combined with the innovations in transdisciplinary approaches, suggesting that proteolytic Abs have opened the way to new translational applications for these tools - the creation of abzymes capable of catalyzing the cleavage of biopolymers is naturally very promising. As such, biocatalysts from the Ig superfamily (Ab-proteases) have significant advantages over conventional proteolytic enzymes.
The interaction between Ags and Abs resembles to some extent that of an enzyme with its substrate; this applies to the key and lock concept as well as to the type of non-covalent interaction. The underlying idea for employing Abs as biocatalysts is that the immunization of an animal with a stable transition state analog of the reaction of interest yields Abs that should be able to catalyze the respective reaction. As a result, catalytic Abs possess unique features capable of both recognizing and enzymatically degrading Ags. In this context, the exquisite specificity, natural biological functions, and favorable development properties of abzymes, and of Ab-proteases, in particular, make them highly effective agents as drugs. Yet beyond this pharmacologic wheelhouse, Ab proteases perform suboptimally for targets of high abundance and those not easily accessible from circulation. Fundamentally, this restraint on broader application is due largely to the stoichiometric nature of their activity-one drug molecule is generally able to inhibit a maximum of two target molecules at a time. Enzymes in contrast are able to catalytically turn-over multiple substrates, making them a natural sub-stoichiometric solution for tar-gets of high abundance or in poorly accessible sites of action. However, enzymes have their own limitations as drugs, including, in particular, the polypharmacology and broad specificity often seen with native enzymes. In this context, Ab-mediated substrate targeting can enhance enzyme activity and specificity, with proof of concept for two challenging target proteins and IgG.
The catalytic activity of an abzyme resides in the V-domain of the Ab, which is constituted by the close spatial arrangement of amino acid residues involved in catalysis. The origin of abzymes is conferred by the innate diversity of the Ig gene repertoire. Un-der immune imbalance, as in autoimmune diseases, the generation of abzymes to autoAgs could be deleterious [46-48]. With advances in engineering technology and the aid of bioinformatics tools, scientists have developed Ab-proteases with unique characteristics and catalytic properties to destroy specific pathogens, remove autoimmune metabolites, and design catalysts to stimulate immunity within the framework of preventive immunization [35,49].
In the work of Daily et al. [50], biocatalysis is considered as processes involving diverse steps of the development of conformational dynamics at different time scales and spatial ones, which are specific of the investigated reactions. If complementarity between the active site and the transition state contributes significantly to enzymatic catalysis, it should be possible to synthesize an abzyme by constructing such an active site. One way to do this is to prepare an Ab to a haptenic group, which resembles the transition state of a given reaction. The combining sites of such Abs should be complementary to the transition state and should cause an acceleration by forcing bound substrates to resemble the transition state. In order for the understanding of the above-mentioned steps, the free-energy panorama of the reaction is to be scanned: as a momentous point, the application of Markov Models has been proposed [50-59].
As we all know, both canonical Abs and enzymes are protein molecules, and the binding of enzymes to substrates and the binding of Abs to Ags are highly specific. But the fundamental difference between these two binding forms is that enzymes bind to high-energy transition-state molecules, while Abs bind to Ags (ground-state molecules). Although their structures are very different, they have two things in common: they are both proteins and they can bind to target molecules with high selectivity. Catalytic Abs have a huge potential for application to many fields including therapeutics. The new technologies based on upgraded algorithm, will contribute hugely to making the desired catalytic Abs without difficulty. Because thousands of original Abs (including mAbs) have been produced, some of them should definitely be converted into the corresponding catalytic Abs including therapeutics.
In modern biotech and daily clinical practice, druggable abzymes (abzymes with therapeutic potential) are selected to bind with high affinity to a target epitope, and thus enzymes based on antibody structure – abzymes-are attractive in that they should have high specificity for their target [60]. These catalytically active Abs are generated by selecting Abs that bind to a stable transition state analogue. Moreover, abzyme-responsive biomaterials are smart materials designed to interact with the body’s natural processes – the evidence-based motivation is driven by the fact that communication between biomaterials and cells is achieved by the development of various materials with enzyme (or abzyme)-responsive moieties in order to respond to cellular cues. In this context, abzyme responsive materials are a class of stimuli responsive materials with broad application potential in biological settings. The strategies employed to incorporate abzyme sensitive functionalities in the biomaterials is often based on common principles, albeit some unique approaches have been developed that merit closer attention [61-71]. For drug design purposes, Ab-proteases can be embedded into biopolymers to create smart materials that respond to environmental changes, such as in drug delivery or tissue regeneration [68-71].
Following the above-mentioned, we would stress the importance of the efforts of introducing Ab-mediated proteolysis to enable selective sub-stoichiometric turnover of therapeutic targets – the Ab-protease-mediated substrate targeting can enhance abzyme activity and specificity, whilst generating a new design-inspired bio therapeutic platform that integrates the favorable properties of Ab-proteases to control therapeutic targets. Attempts are also being made to develop proteolytic Abs containing a catalytic triad analogous to that of serine proteases, designed to cleave gp120 (for treatment of HIV) or IgE (for treatment of allergy) [61,67,68]. In this context, Ab proteases can be programmed and re-programmed to suit the needs of the body metabolism. Or designed for the development of principally new biocatalysts with no natural counterparts (abzymes can be designed to catalyze a wide range of reactions, not performed by natural enzymes, since newest developments in abzymes technology exploit combinatorial libraries). Therefore, the proposed predictive value of Ab-proteases for the development of autoimmune and inflammatory conditions is being challenged and could be utilized in developing diagnostic tools of the next step generation [62].
The field of Ab proteases research is rapidly evolving, with several emerging trends and future directions:
(i) Ab protease-based therapeutics: Are being developed for various diseases, including inflammatory, autoimmune and infectious diseases;
(ii) Ab protease diagnostics: Are being developed as diagnostic and predictive tools PPM-guided clinical practice;
(iii) Ab protease-based biocatalysis: For the production of fine medical chemicals and biopharmaceuticals.
For instance, MPB-, CM- or TG-targeted Ab proteases can be considered to be a promising molecular tool in monitoring patients with autoimmunity diseases at their clinical and subclinical stages. In this context, the primary translational potential of targeted Ab proteases is in the rational design of new therapeutics to exploit the role of the key pathways in influencing disease. By changing sequence specificity, one may reach reduction of a density of the proteolytic effects within the damaged tissue and thus minimizing scales of autoimmune inflammation. Moreover, targeted Ab-mediated proteolysis could be applied to isolate from Ig molecules catalytic domains directed against auto aggressive epitopes or domains containing segments to exert proteolytic activity and then be used as therapeutic modifiers. An important goal in the design of proteolytic Abs is the development of general rules relating happen structure to the corresponding catalytic groups in the Ab combining site [62-64,66].
Conclusion
In addition to classical functions, abzymes exhibit a variety of non-canonical functions related to the destruction of various pathogens and/or oncogenes due to catalytic activity, the action as agonists/antagonists of various receptors, etc. Of special interest is a segment unveiling the secrecy of artificial narcotic-related catalytic Abs (e.g., a rationally designed cocaine antagonist) with potential for medicinal use aiming at eliminating from the human body metabolic waste or providing protection in chemical warfare. The achievements in targeted Ab-mediated catalysis have enriched our deep understanding of life-related biocatalysis, particularly by biomolecules in human biofluids and liquid systems as a whole.
References
- Pauling L (1946) Molecular architecture and biological reactions. Chem Eng News 34: 1375-1377.
- Jencks WP (1969) Catalysis in chemistry and enzymology. McGraw-Hill, New Yok.
- Blackburn GM, Rickard JH, Cesaro-Tadic S, Lagos D, Mekhalfia A, et al. (2004) Passive and catalytic antibodies and drug delivery. Pure Appl Chem 76: 983-989.
- Bowen A, Wear M, Casadevall A (2017) Antibody-mediated catalysis in infection and immunity. Infect Immun 85: e00202-17. [Crossref]
- Zhao D, Chen J, Hu X, Zhang S (2023) Catalytic antibodies: Design, expression, and their applications in medicine. Appl Biochem Biotechnol 195: 1514-1540. [Crossref]
- Sisodiya SM (2021) Precision medicine and therapies of the future. Epilepsia 62: S90-S105. [Crossref]
- Mal'tsev KA, Khitrov AN, Vvedenskaia OIu, Ponomarenko NA, Isaeva MA, et al. (2006) Catalytic autoantibodies--a new molecular instrument in cardiology and ophthalmology. Ter Arkh 78: 70-76. [Crossref]
- Matsuura K, Ohara K, Munakata H, Hifumi E, Uda T (2006) Pathogenicity of catalytic antibodies: Catalytic activity of Bence Jones proteins from myeloma patients with renal impairment can elicit cytotoxic effects. Biol Chem 387: 543-548. [Crossref]
- Jones LH, Wentworth P Jr (2001) The therapeutic potential for catalytic antibodies: From a concept to a promise. Mini Rev Med Chem 1: 125-132. [Crossref]
- Amital H, Tur-Kaspa I, Tashma Z, Hendler I, Shoenfeld Y (1995) Catalytic antibodies. Structure and possible applications. Methods Mol Biol 51: 203-210. [Crossref]
- Ramesh R, Sundaresh A, Rajkumar RP, Negi VS, Vijayalakshmi MA, et al. (2021) DNA hydrolysing IgG catalytic antibodies: An emerging link between psychoses and autoimmunity. NPJ Schizophr 7: 13. [Crossref]
- Jin Y, Huang Z, Xu B, Chen J (2023) Localization of multiple DNAzymes as a branchedzyme-powered nanodevice for the immunoassay of tumor biomarkers. Anal Chim Acta 1274: 341580. [Crossref]
- Wootla B, Lacroix-Desmazes S, Warrington AE, Bieber AJ, Kaveri SV, et al. (2011) Autoantibodies with enzymatic properties in human autoimmune diseases. J Autoimmun 37: 144-150.
- Li L, Kaveri S, Tyutyulkova S, Kazatchkine MD, Paul S (1995) Catalytic antibodies in human immunity. Ann N Y Acad Sci 764: 570-572.
- Suchkov S, Schroeder H, Rose N, Gabibov A (2018) Antibody-proteases as highly informative biomarkers and efficient targets of the newest generation. Meta Gene 17: S18-S19.
- Isaeva MA, Bogatyreva ZI, Andreeva A, Suchkova EN, Korotkova AA, et al. (2010) Current protease-antibody-based serodiagnostic tools and their use in the diagnosis of autoimmune thyroid diseases. Klin Lab Diagn 4: 37-42.
- Wootla B, Dasgupta S, Dimitrov JD, Bayry J, Levesque H, et al. (2008) Factor VIII hydrolysis mediated by anti-factor VIII autoantibodies in acquired hemophilia. J Immunol 180: 7714-7720. [Crossref]
- Black RS, Sperling RA, Safirstein B, Motter RN, Pallay A, et al. (2010) A single ascending dose study of bapineuzumab in patients with Alzheimer disease. Alzheimer Dis Assoc Disord 24: 198-203. [Crossref]
- Taguchi H, Planque S, Nishiyama Y, Symersky J, Boivin S, et al. (2008) Autoantibody-catalyzed hydrolysis of amyloid beta peptide. J Biol Chem 283: 4714-4722. [Crossref]
- Song C, Li H, Zhang T, Zheng C, Zhang Y (2024) Synergetic effect of matrine on the catalytic scFv antibody HS72 in vitro and in mice with Alzheimer disease pathology. Neuropharmacology 242: 109775. [Crossref]
- Ponomarenko NA, Durova OM, Vorobiev II, Belogurov AA, Telegin GB, et al. (2006) Catalytic activity of autoantibodies toward myelin basic protein correlates with the scores on the multiple sclerosis expanded disability status scale. Immunol Lett 103: 45-50. [Crossref]
- Paul A, Comabella M, Gandhi R (2019) Biomarkers in multiple sclerosis. Cold Spring Harb Perspect Med 9: 1-22. [Crossref]
- Polosukhina DI, Kanyshkova TG, Doronin BM, Tyshkevich OB, Buneva VN, et al. (2004) Hydrolysis of myelin basic protein by polyclonal catalytic IgGs from the sera of patients with multiple sclerosis. J Cell Mol Med 8: 359-368. [Crossref]
- Ponomarenko NA, Durova OM, Vorobiev II, Aleksandrova ES, Telegin GB, et al. (2002) Catalytic antibodies in clinical and experimental pathology: Human and mouse models. J Immunol Methods 269: 197-211. [Crossref]
- Nevinsky GA, Baranova SV, Buneva VN, Dmitrenok PS (2021) Multiple sclerosis: Enzymatic cross site-specific hydrolysis of H1 histone by IgGs against H1, H2A, H2B, H3, H4 histones, and myelin basic protein. Biomolecules 11: 1140.
- Belogurov AA, Kurkova IN, Friboulet A (2008) Recognition and degradation of myelin basic protein peptides by serum autoantibodies: Novel biomarker for multiple sclerosis. J Immunol 180: 1258-1267.
- Belogurov AA, Kurkova IN, Misikov VK, Suchkov SV, Telegin GB, et al. (2007) Substrate specificity of catalytic autoantibodies in neurodegenerative processes. Dokl Biochem Biophys 413: 61-64. [Crossref]
- Medveczky P, Antal J, Patthy A, Kékesi K, Juhász G, et al. (2006) Myelin basic protein, an autoantigen in multiple sclerosis, is selectively processed by human trypsin 4. FEBS Lett 580: 545-552. [Crossref]
- Scarisbrick IA (2008) The multiple sclerosis degradome: Enzymatic cascades in development and progression of central nervous system inflammatory disease. Curr Top Microbiol Immunol 318: 133-175. [Crossref]
- Kamaeva DA, Smirnova LP, Vasilieva SN, Kazantseva DV, Vasilieva AR, et al. (2022) Catalytic antibodies in bipolar disorder: Serum IgGs hydrolyze myelin basic protein. Int J Mol Sci 23: 7397.
- Jensen MA, Dafoe ML, Wilhelmy J, Cervantes L, Okumu AN, et al. (2024) Catalytic antibodies may contribute to demyelination in myalgic encephalomyelitis/chronic fatigue syndrome. Biochem 63: 9-18. [Crossref]
- Gabibov AG, Paltsev MA, Suchkov SV (2011) Antibody-associated proteolysis in surveillance of autoimmune demyelination: Clinical and preclinical issues. Future Neurol 6: 303-305.
- Kostyushev D, Tsarev I, Gnatenko D, Paltsev M, Suchkov S, et al. (2011) Myelin-associated serological targets as applicable to diagnostic tools to be used at the preclinical and transient stages of multiple sclerosis progression. Open J Immunol 1: 80-86.
- Lacroix-Desmazes S, Bayry J, Kaveri SV, Hayon-Sonsino D, Thorenoor N, et al. (2005) High levels of catalytic antibodies correlate with favorable outcome in sepsis. Proc Natl Acad Sci USA 102: 4109-4113. [Crossref]
- Mahendra A, Sharma M, Rao DN, Peyron I, Planchais C, et al. (2013) Antibody-mediated catalysis: Induction and therapeutic relevance. Autoimmun Rev 12: 648-652. [Crossref]
- Geanes ES, McLennan R, LeMaster C, Bradley T (2024) Autoantibodies to ACE2 and immune molecules are associated with COVID-19 disease severity. Commun Med (Lond) 4: 47. [Crossref]
- Bogatyreva ZI, Suchkova EN, Isaeva MA, Unanyan VN, Notkins AL, et al. (2014) Antithyroid autoantibody population as a source of antibody specificity and functionality: Clinical significance of the phenomenon of the serological orchestra in monitoring of patients with Graves' disease. EPMA J 5: A142.
- Leuschner F, Katus HA, Kaya Z (2009) Autoimmune myocarditis: Past, present and future. J Autoimmun 33: 282-289. [Crossref]
- Caforio AL, Tona F, Bottaro S, Vinci A, Dequal G, et al. (2008) Clinical implications of anti-heart autoantibodies in myocarditis and dilated cardiomyopathy. Autoimmunity 41: 35-45. [Crossref]
- Sun M, Gao QS, Kirnarskiy L, Rees A, Paul S, et al. (1997) Cleavage specificity of a proteolytic antibody light chain and effects of the heavy chain variable domain. J Mol Biol 271: 374-385.
- Suchkov SV (2020) Antibodies with functionality as a new generation of translational tools designed to be programmed via translational research. IOMC World.
- Polosukhina DI, Kanyshkova TG, Doronin BM, Tyshkevich OB, Buneva VN, et al. (2004) Hydrolysis of myelin basic protein by polyclonal catalytic IgGs from the sera of patients with multiple sclerosis. J Cell Mol Med 8: 359-368. [Crossref]
- Belogurov AA, Kurkova IN, Misikov VK, Suchkov SV, Telegin GB, et al. (2007) Substrate specificity of catalytic autoantibodies in neurodegenerative processes. Dokl Biochem Biophys 413: 61-64.
- Erster O, Thomas J, Hamzah J, Jabaiah A, Getz J, et al. (2012) Site-specific targeting of antibody activity in vivo mediated by disease-associated proteases. J Control Release 161: 804-812. [Crossref]
- Ermakov EA, Nevinsky GA, Buneva VN (2020) Immunoglobulins with non-canonical functions in inflammatory and autoimmune disease states. Int J Mol Sci 21: 5392. [Crossref]
- Avalle B, Friboulet A, Thomas D (2000) Enzymes and abzymes relationships. J Mol Catal B Enzym 10: 39-45.
- Thomas D (2000) Enzymes and abzymes Relationships. J Mol Catal B Enzym 10: 39-45.
- Hilvert D (1991) Extending the chemistry of enzymes and abzymes. Trends Biotechnol 9: 11-17.
- Padiolleau-Lefèvre S, Ben Naya R, Shahsavarian MA, Friboulet A, Avalle B, et al. (2014) Catalytic antibodies and their applications in biotechnology: State of the art. Biotechnol Lett 36: 1369-1379. [Crossref]
- Daily MD, Yu H, Phillips GN Jr, Cui Q (2013) Allosteric activation transitions in enzymes and biomolecular motors: Insights from atomistic and coarse-grained simulations. Top Curr Chem 337: 139-164. [Crossref]
- Noé F, Schütte C, Vanden-Eijnden E, Reich L, Weikl TR. Constructing the equilibrium ensemble of folding pathways from short off-equilibrium simulations. Proc Natl Acad Sci 106: 19011-19016. [Crossref]
- Lecian OM (2003) Analytical results from the two-state Markovv-states model and applications to validation of molecular dynamics. IJMCR 11: 3746- 3754.
- Tozzini V (2010) Minimalist models for proteins: A comparative analysis. Q Rev Biophys 43: 333-371. [Crossref]
- Bowman GR, Beauchamp KA, Boxer G, Pande VS, Bowman GR, et al. (2009) Progress and challenges in the automated construction of Markov state models for full protein systems. Journ Chem Phys 131: 124101 [Crossref]
- Vanden-Eijnden EWE (2006) Toward a theory of transition paths. J Stat Phys 123: 503-523.
- Metzner P, Schuette C, Vanden-Eijnden E (2009) Transition path theory for Markov jump processes. Multiscale Model Simul 7: 1192-1219.
- Du R, Pande VS, Alexander, Anaka TT, Shakhnovich ES (1998) On the transition coordinate for protein folding. J Chem Phys 108: 334-350.
- Bolhuis PG, Chandler D, Dellago C, Geissler PL (2002) Transition path sampling: Throwing ropes over rough mountain passes, in the dark. Annu Rev Phys Chem 53: 291-318. [Crossref]
- Lecian OM (2025) Markov chains of irreversible reactions, Researchgate, Berlin, Germany.
- Rational design of an abzyme (1998) Nat Biotechnol Rational design of an abzyme.
- Drug Discovery and Development (2012) Technology in Transition. 2nd Edition. Churchill Livingstone. Editor: Raymond G Hill.
- Romei MG, Leonard B, Kim I, Kim HS, Lazar GA (2023) Antibody-guided proteases enable selective and catalytic degradation of challenging therapeutic targets. J Biol Chem 299: 104685.
- Wenthur CJ, Cai X, Ellis BA, Janda KD (2017) Augmenting the efficacy of anti-cocaine catalytic antibodies through chimeric hapten design and combinatorial vaccination. Bioorg Med Chem Lett 27: 3666-3668. [Crossref]
- Wang Y, Rickhaus M, Blacque O, Baldridge KK, Juríček M, et al. (2022) Cooperative weak dispersive interactions actuate catalysis in a shape-selective abiological racemase. J Am Chem Soc 144: 2679-2684. [Crossref]
- Emi Hifumi, Hiroaki Taguchi, Tamami Nonaka, Taizo Uda (2023) Direct conversion of a general anti-body to its catalytic antibody and corresponding applications -Importance and role of Pro95 in CDR-3. Proc Jpn Acad Ser B Phys Biol Sci 99: 155-172. [Crossref]
- Uda T, Kato R, Shigeta Y, Hirota S, Kobayashi J, et al. (2025) Structural and biochemical differences between non-catalytic and catalytic antibodies. MAbs 17: 2503978. [Crossref]
- Mei M, Tan X (2021) Current strategies of antiviral drug discovery for COVID-19. Front Mol Biosci 8: 671263. [Crossref]
- Yang Z, Lushington GH (2025) Exploring the blueprint of life: The innovation in antibody and protein design. Comb Chem High Throughput Screen 29: 551-554. [Crossref]
- Brouns JEP, Dankers PYW (2021) Introduction of enzyme-responsivity in biomaterials to achieve dynamic reciprocity in cell-material interactions. Biomacromolecules 22: 4-23. [Crossref]
- Pi Y, Ganabady K, Celiz AD (2025) Enzyme-responsive biomaterials for biomedical applications. Commun Mater 6: 263.
- Urusov AE, Tolmacheva AS, Aulova KS, Nevinsky GA (2023) Autoantibody–abzymes with catalase activity in experimental autoimmune encephalomyelitis mice. Molecules 28: 1330. [Crossref]