Free-form amino acids were quantitatively measured in over 30 species of vegetables and mushrooms. Significant amounts of free-form amino acids were found in each species analyzed. Additionally, there appear to be no rules or patterns of free-form amino acid distribution among species. Highly functional free-form amino acids, like γ-aminobutyric acid, are found in significant levels and suggest that amino acid analysis could play an informative role in nutrition. Our analysis should provide valuable data for the establishment of nutritional databases that include free-form amino acids and more accurately represent the entire amino acid profile of foods.
free-form amino acid, -aminobutyric acid, mushroom, supplements, vegetable
Amino acids play significant biological roles in human. The amino acids required for human metabolism arise from the digestion and utilization of amino acids from food, and from free-form amino acids within food components (For a list of amino acid abbreviations used in this study: Table 1). Free-form amino acids may be utilized as nutrients and/or act as messengers that target specific receptors.
Table 1. List of amino acids and their abbreviations
Amino acid |
3 Letter abbreviation |
Alanine |
Ala |
β-Alanine |
β-Ala |
Arginine |
Arg |
Asparagine |
Asn |
Aspartic acid |
Asp |
Citrulline |
Cit |
Cysteine |
Cys |
Cystine |
Cys-Cys |
γ-Amino butyric acid |
GABA |
Glutamic acid |
Glu |
Glutamine |
Gln |
Glycine |
Gly |
Histidine |
His |
Hydroxyproline |
HYP |
Isoleucine |
Ile |
Leucine |
Leu |
Lysine |
Lys |
Methionine |
Met |
Ornithine |
Orn |
Phenyl alanine |
Phe |
Proline |
Pro |
Serine |
Ser |
Threonine |
Thr |
Tryptophan |
Trp |
Tyrosine |
Tyr |
Valine |
Val |
Free-form amino acids exist in foods derived from plants and animals. β-alanine (β-Ala), citrulline (Cit), g-aminobutyric acid (GABA), and ornithine (Orn) usually exist as free-form amino acids and perform various biological functions including coenzyme components, control of osmoregulation, ammonia detoxification, and as neurotransmitters [1,2]. In human, tryptophan (Trp), a precursor of serotonin and melatonin, acts to regulate circadian rhythm [3]. Some free-form amino acids act as neurotransmitters: in the central nervous system GABA and glycine (Gly) function as inhibitory signals while glutamic acid (Glu) is an excitatory signal [4-8] and may have similar functions in non-neuronal cells [5]. The roles of free-form amino acids in dietary supplements have been extensively explored and dietary branched chain amino acids (BCAA; including valine (Val), leucine (Leu), and isoleucine (Ile)), improve exercise performance, endurance performance, and upper body power in human [9]. Moreover, GABA can reduce high blood pressure [10], and in Japan is authorized as “Tokuho”, which is a “food for specified health uses”, and other health foods containing GABA are widely available.
Glu receptors have been identified on taste buds and the gastrointestinal tract, including stomach cells [11], which suggests that free-form Glu may be involved in enhancing digestion, nutrient absorption, and/or utilization of nutrients [12]. The study was extended to examine the use of daily dietary Glu supplements in the meals of elderly people in hospital [12]. The results of this analysis showed that Glu supplementation improved the nutritional status of elderly and nutritionally deficient patients through improved food intake [12-14]. Therefore, free-form Glu is a highly effective supplement with the potential to improve quality of life (QOL) [12,15,16]. It is likely that free-form amino acids exert useful effects on physiologic functions, particularly during food digestion and nutrient absorption.
Some free-form amino acids act as palatable compounds and are key determinants in the taste of food. Free-form Glu, also known as monosodium glutamate or MSG, is an umami compound isolated from kelp, and is also rich in tomatoes and cheese [16]. Other free-form amino acids that confer taste include Gly, alanine (Ala), and proline (Pro) which provide sweetness, and Leu, Ile, and Val which provide a bitter taste [17,18]. L-serine (Ser) gives mainly sweetness and minor umami taste, and D-Ser, enantiomer of L-Ser, has sweetness. As for taste of D-amino acids, it is known that D-Ala and D-Leu are sweeter than their L-enantiomers [19]. Amino acids elicit not only single basic tastes but also complex taste when they are combined.
Information about the "amino acid" content of various agricultural products is available in databases, typically called "Standard Tables of Food Composition". However, amino acid databases usually list amino acid composition measured in acid hydrolyzed samples, so most of these databases do not accurately reflect free-form amino acid contents [20]. The Stein-Moore laboratory, at Rockefeller University, has shown that acid hydrolysis destroys glutamine (Gln), asparagine (Asn), Trp, and some other amino acids such as GABA, Orn, Cit [21,22]. Thus, little is known about the kinds and amounts of free-form amino acids present in foods.
Studies of nutrition, particularly concerning the quality of proteins, have progressed extensively since information about the amino acid composition of foods has become readily available. However, information about the amino acid composition of both hydrolyzed samples and free-form amino acids in the original food samples are necessary for analyses of palatability and the beneficial functions of amino acids. The Japan Society of Nutrition and Food Science website lists some information about free-form amino acids in food, but the data presented was collated from literature published about 30 years ago [23-26]. Therefore, the amino acid contents listed in some of these databases are likely to be misleading or incomplete.
Amino acid analysis has advanced over the last few decades, and the time has come to renew these data using modern techniques that provide increased sensitivity and reliability. Currently, only a few databases specializing in free-form amino acids are available. To understand the amount of free-form amino acids in our daily foods, free-form amino acids must be "extracted" and then measured. Previously, we reported initial attempts to establish a free-form amino acid database for approximately 140 species of fruits and vegetables [27]. We showed that distribution of free-form amino acids is characteristic for each sample and significantly different from the amino acid distribution recorded in acid hydrolyzed databases. Here, we have extended our analysis of free-form amino acid contents to include additional vegetables and some mushroom species.
Extraction of free amino acids from foods
We examined 38 species of vegetables and mushrooms (Table 2). Some samples were purchased from local supermarkets in Shiga Prefecture and others were supplied by local farmers. The food samples came from various geographic regions. All food samples were washed in water and the edible and inedible parts, including peels, seeds, and skins, were separated. Samples were stored at -25°C until use.
Table 2. List of foods analyzed for free-form amino acids
|
Scientific Name |
Food |
Vegetables |
Abelmoschus esculentus |
Okra “Yashiro-shio okura” |
Allium cepa |
Onion |
Allium cepa |
Red onion |
Capsicum annuum |
Paprika |
Chrysanthemum morifolium |
Edible chrysanthemum |
Chrysanthemum morifolium |
Edible purple chrysanthemum “Enmeiraku” |
Cucurbita maxima |
Winter squash |
Cucurbita moschata |
Japanese squash |
Daucus carota |
young carrot (Leaf) |
Daucus carota |
Moso bamboo |
Phyllostachys heterocycla |
Pea |
Pisum sativum |
Peas (Tutankhamen) |
Pisum sativum |
Podded pea |
Raphanus sativus |
Radish sprout |
Rheum rhabarbarum |
Garden rhubarb |
Solanum mlycopersicum |
Cherry tomato |
Solanum melongena |
Egg plant “Beinasu” |
Solanum melongena |
Egg plant “Kamonasu” |
Solanum melongena |
Egg plant “Kyokonasu” |
Solanum melongena |
Egg plant “Marunasu” |
Solanum melongena |
Egg plant “Zebranasu” |
Spinacia olerace |
Spinach “Okame” |
Vicia faba |
Broad bean |
Vigna unguiculata |
Cowpea |
Vigna unguiculata
|
Cowpea beans |
Pumpkin “Korinnki” |
Mushrooms |
Agaricus bisporus |
Button mushroom (brown) |
Agaricus bisporus |
Button mushroom (white) |
Auricularia auricula-judae |
Jew’s ear |
Boletus edulis |
Porcini mushrooms (Dried) |
Flammulina velutipes |
Winter mushroom |
Grifola frondosa |
Mushroom “Maitake” |
Hypsizygus marmoreus |
Beech mushroom |
Hypsizygus marmoreus |
Beech mushroom(Tanba) |
Letinula edodes |
Mushroom “Shiitake” |
Lyophyllum shimeji |
Mushroom “Honshimeji” |
Pleurotus eryngii |
King oyster mushroom |
Sample preparation
Samples (5-10 g) were ground to a fine powder with a mortar and pestle. The powdered samples were added to 100 mM HEPES-Na buffer, pH 7.0, and homogenized on ice with a Tissue-Tearor (Biospec). The homogenate was centrifuged at 10,000×g for 10 min and the supernatant was collected. Suitable supernatant aliquots were treated with 60% perchloric acid to remove proteins. The protein content of samples was estimated by Bradford protein assays (BioRad). Ultrafiltration was performed using CENTRICON®10 (Millipore) when further removal of protein contamination was necessary. After ultrafiltration, the protein content was measured by Bradford protein assay.
Amino acid standards
GABA, hydroxyproline (HYP), L-Gln, L-Asn, β-Ala, L-cysteine (Cys), and L-Trp were purchased from Wako Chemicals. Purchased amino acids were added to type H amino acid mixture standard solution (Wako Chemicals), to produce a working standard solution of each amino acid (100 µmol/L in 0.1 N HCl).
Amino acid analysis
Free-form amino acids extracted from food samples were analyzed on a Hitachi Ultra High Speed Liquid Chromatography system equipped with UV-Vis detector. Samples were reacted with 4-fluoro-7-nitrobenzofurazan (NBD-F) and derivatized NBD-amino acids were separated on a Zorbax SB-C18 column (3.0 × 50 mm, 2.7 µm; Agilent Technologies) and elution was monitored at 470 nm. Derivatization of amino acids was performed according to the Hitachi Ultra High Speed Liquid Chromatography manual. NBD-amino acids were eluted using a stepwise gradient of increasing B solution: equilibration with 15%, 15-25% for 2.7 min, 25-35% for 3.5 min, 35-70% for 1.0 min, 70-85% for 0.1 min, and kept at 85% for 1.5 min, in sequence. Elution solutions A and B were purchased from Hitachi High-Technologies, where solution A mainly consists of ammonium formate and solution B of acetonitrile. The flow rate was maintained at 0.550 mL/min throughout the analysis.
Linearity of the dose response was evaluated using an amino acid standard solution within the range of 10 to 60 pmol. Minimum and maximum correlation coefficients (r2) for Asn, as an example, were 0.974 and 0.997, respectively. Ten consecutive standard solution injections were analyzed to evaluate reproducibility (Table 3). The relative standard deviation of peak area for Tyr and His was in the range of 0.3% and 5.1%, respectively. The mean limit of detection was 0.8 pmol.
Table 3. Means of peak area and retention time (min) 10 replicates analysis
|
Peak Area |
Elution Time (min) |
Mean ± SE |
%RSD |
Mean ± SE |
%RSD |
His |
107839 ± 1724 |
5.05 |
0.784 ± 0.001 |
0.290 |
Arg |
100894 ± 1427 |
4.47 |
0.909 ± 0.004 |
1.250 |
HYP |
113930 ± 589 |
1.64 |
1.715 ± 0.004 |
0.740 |
Asn |
34137 ± 497 |
4.61 |
2.003 ± 0.001 |
0.216 |
Gln |
50739 ± 470 |
2.93 |
2.158 ± 0.002 |
0.224 |
Cit |
79837 ± 342 |
1.36 |
2.250 ± 0.001 |
0.166 |
Ser |
53349 ± 478 |
2.83 |
2.331 ± 0.002 |
0.301 |
Asp |
48268 ± 530 |
3.47 |
2.760 ± 0.001 |
0.156 |
Glu |
51824 ± 418 |
2.55 |
2.927 ± 0.001 |
0.101 |
Thr |
107560 ± 300 |
0.88 |
3.073 ± 0.001 |
0.094 |
Pro+β-Ala |
106552 ± 302 |
0.90 |
3.156 ± 0.001 |
0.132 |
Gly |
90638 ± 637 |
2.22 |
3.254 ± 0.002 |
0.166 |
GABA |
62614 ± 510 |
2.57 |
3.551 ± 0.002 |
0.141 |
Ala |
53683 ± 593 |
3.49 |
3.817 ± 0.001 |
0.124 |
Val |
78636 ± 237 |
0.95 |
5.465 ± 0.002 |
0.139 |
Met |
61485 ± 95 |
0.49 |
5.612 ± 0.001 |
0.080 |
Leu |
68875 ± 316 |
1.45 |
6.363 ± 0.002 |
0.099 |
Ile |
84374 ± 233 |
0.87 |
6.519 ± 0.003 |
0.140 |
Cys-Cys |
56979 ± 197 |
1.09 |
6.689 ± 0.002 |
0.118 |
Trp |
69741 ± 295 |
1.34 |
6.820 ± 0.002 |
0.078 |
Orn |
74663 ± 277 |
1.17 |
6.864 ± 0.002 |
0.089 |
Phe |
80193 ± 134 |
0.53 |
6.947 ± 0.002 |
0.069 |
Lys |
79550 ± 210 |
0.83 |
7.026 ± 0.001 |
0.063 |
Cys |
82425 ± 178 |
0.68 |
7.226 ± 0.001 |
0.029 |
Tyr |
61109 ± 62 |
0.32 |
7.450 ± 0.001 |
0.031 |
Amino acids are listed as abbreviated according to Table 1.
Chromatograms for the amino acid mixture standard and eggplant "kamonasu" (Solanum melongena) were obtained and analyzed (Figures 1 and 2). In our system, the typical time to baseline NBD-amino acid separation, with the exception of Pro and β-Ala, was about 8 min.
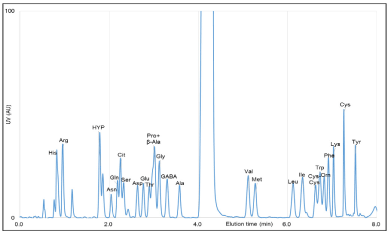
Figure 1. HPLC chromatogram of the amino acids standard mixture derivatized with NBD-F and UV-VIS detection
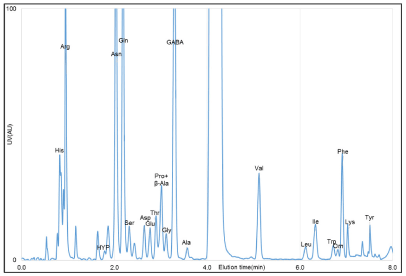
Figure 2. HPLC chromatogram of the “kamonasu” derivatized with NBD-F and UV-VIS detection
The free-form amino acid contents of the vegetables and mushrooms analyzed varied considerably (Tables 4 and 5). The total free-form amino acids found in the extracts ranged from 59.91 (Garden rhubarb) to 4,093 and 424.6 (Jew's ear) to 12,773 µmol per 100 g of vegetables and mushrooms, respectively. Of the vegetables and mushrooms analyzed, the podded pea (4,093 µmol per 100 g) and dried porcini mushrooms (12,773 µmol per 100 g) had the highest total free-form amino acids of vegetables and mushrooms, respectively. The most abundant free-form amino acids found in the vegetables examined were Asn, Gln, Glu, and GABA, all of which are important metabolites in human. In mushrooms, arginine (Arg), Glu, Gln, and Ala were the most abundant free-form amino acids. Orn was found in mushrooms, but rarely in vegetables and HYP, Cys, and methionine (Met) were found at low levels in both vegetables and mushrooms.
Table 4. Contents of free-form amino acids assayed in vegetables (µmol/100 g sample on a wet weight basis)
|
Okra"Yashiro-shio okura" |
Onion |
Red onion |
Paprika |
Edible chrysanthemum
"Kinkaramatsu" |
Edible chrysanthemum
"Enmeiraku" |
His |
39.5 ± 0.977 |
16.06 ± 4.234 |
8.394 ± 0.674 |
10.68 ± 1.84 |
13.44 ± 3.137 |
9.415 ± 1.225 |
Arg |
86.12 ± 2.473 |
83.6 ± 5.363 |
53.49 ± 4.528 |
44.01 ± 2.39 |
32.75 ± 7.504 |
17.23 ± 3.145 |
HYP |
nd |
nd |
nd |
0.32 ± 0.261 |
nd |
nd |
Asn |
1480 ± 40.59 |
79.14 ± 8.421 |
87.73 ± 4.638 |
473.1 ± 45.43 |
408.1 ± 116 |
128.1 ± 7.235 |
Gln |
908 ± 2.846 |
157.3 ± 10.06 |
286.2 ± 19.35 |
255.8 ± 23.08 |
469.3 ± 111.9 |
596.7 ± 51.47 |
Cit |
nd |
nd |
nd |
nd |
nd |
nd |
Ser |
129.1 ± 3.901 |
15.97 ± 1.216 |
24.37 ± 1.356 |
99.13 ± 12.57 |
53.32 ± 14.24 |
61.87 ± 4.289 |
Asp |
46.51 ± 0 |
15.3 ± 1.811 |
16.42 ± 0.921 |
83.82 ± 15.04 |
76.64 ± 14.9 |
115.3 ± 11.64 |
Glu |
105.3 ± 5.084 |
29.89 ± 1.902 |
29.09 ± 1.11 |
36.91 ± 4.401 |
98.7 ± 19.51 |
143.7 ± 9.575 |
Thr |
42.67 ± 1.133 |
135.9 ± 9.568 |
95.39 ± 5.964 |
36.7 ± 7.325 |
58.42 ± 15.05 |
55.44 ± 3.696 |
Pro+β-Ala |
18.15 ± 0.806 |
4.231 ± 0.167 |
6.233 ± 0.567 |
19.2 ± 3.532 |
103.1 ± 17.66 |
24.09 ± 1.438 |
Gly |
22.59 ± 0.228 |
4.777 ± 0.204 |
3.875 ± 0.332 |
17.28 ± 5.297 |
6.604 ± 3.816 |
7.871 ± 0.686 |
GABA |
79.61 ± 11.49 |
4.982 ± 0.523 |
2.74 ± 0.237 |
36.21 ± 4.637 |
0.831 ± 0.831 |
nd |
Ala |
53.62 ± 3.453 |
14.66 ± 1.369 |
22.93 ± 1.376 |
21.45 ± 2.529 |
17.69 ± 5.758 |
33.81 ± 3.291 |
Val |
35.36 ± 2.323 |
19.74 ± 1.286 |
19.77 ± 0.994 |
40.05 ± 7.5 |
26.74 ± 6.647 |
23.73 ± 2.163 |
Met |
nd |
nd |
nd |
4.971 ± 0.58 |
nd |
nd |
Leu |
21.44 ± 0 |
28.89 ± 1.684 |
31.6 ± 1.652 |
15.5 ± 1.354 |
5.079 ± 2.957 |
nd |
Ile |
13.34 ± 0.802 |
15.14 ± 0.701 |
13.66 ± 1.315 |
9.44 ± 0.669 |
14.58 ± 5.23 |
6.451 ± 0.718 |
Cys-Cys |
nd |
nd |
nd |
nd |
nd |
nd |
Trp |
16.61 ± 0.042 |
9.72 ± 1.496 |
6.809 ± 0.757 |
nd |
3.274 ± 1.375 |
nd |
Orn |
nd |
nd |
nd |
nd |
nd |
nd |
Phe |
44.37 ± 0.51 |
21.26 ± 1.237 |
20.46 ± 1.604 |
9.36 ± 0.673 |
21.3 ± 5.56 |
4.035 ± 0.349 |
Lys |
4.88 ± 0.641 |
25.19 ± 3.425 |
27.54 ± 2.864 |
4.736 ± 2.777 |
0.456 ± 0.456 |
4.164 ± 0.705 |
Cys |
0.442 ± 0.312 |
nd |
nd |
25.03 ± 5.24 |
0.647 ± 0.373 |
nd |
Tyr |
23.56 ± 0.98 |
31.61 ± 1.768 |
20.01 ± 0.273 |
11.63 ± 0.219 |
4.63 ± 1.658 |
nd |
|
Winter squash |
Japanese squash |
Corinky squash |
Young carrot (Root) |
Young carrot (Leaf) |
Moso bamboo |
His |
74.06 ± 1.997 |
57.78 ± 1.118 |
77.19 ± 4.327 |
0.665 ± 0.47 |
7.221 ± 0.163 |
36.11 ± 2.543 |
Arg |
37.9 ± 10.38 |
15.37 ± 1.592 |
26.6 ± 3.459 |
1.141 ± 0.807 |
0.485 ± 0.343 |
52.1 ± 5.874 |
HYP |
1.134 ± 0 |
nd |
nd |
nd |
nd |
nd |
Asn |
82.25 ± 2.595 |
1025 ± 37.8 |
162.1 ± 16.04 |
2.948 ± 0.458 |
17.41 ± 1.325 |
995.7 ± 151.2 |
Gln |
90.75 ± 6.974 |
367.3 ± 14.21 |
281.9 ± 35.3 |
16.13 ± 5.61 |
55.48 ± 6.236 |
368.6 ± 68.51 |
Cit |
nd |
nd |
nd |
nd |
nd |
nd |
Ser |
63.67 ± 3.586 |
231.2 ± 15.89 |
90.86 ± 9.239 |
11.83 ± 0.405 |
24.09 ± 1.003 |
232.7 ± 42.35 |
Asp |
192.5 ± 15.57 |
118.9 ± 5.924 |
179.7 ± 17.7 |
18.45 ± 4.924 |
51.98 ± 3.635 |
101.7 ± 10.36 |
Glu |
97.7 ± 4.456 |
95.33 ± 14.87 |
21.5 ± 2.688 |
14.09 ± 0.914 |
59.27 ± 5.786 |
63.96 ± 6.861 |
Thr |
23.66 ± 2.029 |
46.92 ± 2.309 |
44.8 ± 5.556 |
7.677 ± 0.336 |
24.43 ± 2.651 |
103.1 ± 13.78 |
Pro+β-Ala |
14.37 ± 1.468 |
59.87 ± 1.693 |
15.03 ± 1.776 |
1.222 ± 0.096 |
8.544 ± 1.058 |
132.6 ± 26.76 |
Gly |
8.521 ± 0.167 |
34.47 ± 1.265 |
9.774 ± 1.11 |
0.286 ± 0.202 |
2.966 ± 0.239 |
32.78 ± 4.574 |
GABA |
112.2 ± 10.17 |
82.02 ± 3.727 |
70.72 ± 6.325 |
40.13 ± 6.319 |
40.4 ± 1.376 |
75.83 ± 12.81 |
Ala |
43.4 ± 1.337 |
181.8 ± 9.326 |
50.15 ± 4.89 |
9.717 ± 2.299 |
49.69 ± 4.37 |
128.9 ± 31.91 |
Val |
14.99 ± 1.734 |
55.74 ± 2.064 |
56.93 ± 5.973 |
5.45 ± 0.333 |
21.05 ± 2.243 |
140.8 ± 26.69 |
Met |
7.215 ± 1.251 |
26.21 ± 1.322 |
20.58 ± 2.241 |
nd |
nd |
44.02 ± 5.688 |
Leu |
40.01 ± 1.073 |
42.37 ± 5.847 |
54.94 ± 6 |
1.852 ± 0.413 |
5.699 ± 0.683 |
110.6 ± 11.53 |
Ile |
15.85 ± 2.566 |
28.14 ± 0.981 |
35.2 ± 3.677 |
2.252 ± 0.407 |
10.44 ± 1.489 |
88.01 ± 12.15 |
Cys-Cys |
nd |
nd |
nd |
nd |
nd |
nd |
Trp |
4.827 ± 0.255 |
20.78 ± 1.077 |
5.034 ± 0.615 |
nd |
nd |
39.6 ± 2.042 |
Orn |
nd |
nd |
nd |
nd |
nd |
nd |
Phe |
29.65 ± 1.843 |
30.89 ± 1.656 |
41.53 ± 4.733 |
2.879 ± 0.04 |
5.631 ± 0.047 |
55.34 ± 5.782 |
Lys |
2.929 ± 0.313 |
1.216 ± 0 |
14.8 ± 2.154 |
1.134 ± 0.802 |
4.007 ± 0.486 |
23.13 ± 2.955 |
Cys |
0.476 ± 0 |
nd |
nd |
nd |
nd |
1.128 ± 0.142 |
Tyr |
34.79 ± 4.246 |
54.89 ± 3.071 |
44.66 ± 5.257 |
2.195 ± 0.367 |
4.417 ± 0.317 |
361.9 ± 32.95 |
|
Pea |
Peas “Tutankhamen” |
Podded pea |
Radish sprout |
Garden rhubarb |
|
His |
49.61 ± 9.239 |
28.71 ± 1.118 |
54.13 ± 4.181 |
70.19 ± 12.28 |
0.598 ± 0.226 |
Arg |
1304 ± 77.94 |
703.6 ± 38.32 |
70.3 ± 12.99 |
33.05 ± 7.199 |
1.391 ± 0.452 |
HYP |
nd |
nd |
nd |
1.762 ± 1.205 |
nd |
Asn |
291 ± 60.56 |
171.5 ± 12.55 |
1257 ± 61.92 |
60.09 ± 13.23 |
7.21 ± 1.421 |
Gln |
688.1 ± 50.39 |
546 ± 60.41 |
890.1 ± 7.863 |
438.7 ± 77.95 |
16.42 ± 1.526 |
Cit |
nd |
nd |
nd |
nd |
nd |
Ser |
153.8 ± 11.01 |
118.1 ± 9.087 |
444.1 ± 24.78 |
60.98 ± 12.41 |
3.174 ± 0.397 |
Asp |
161.4 ± 10.26 |
53.84 ± 7.253 |
114.5 ± 0.625 |
5.169 ± 1.378 |
4.756 ± 0.497 |
Glu |
257 ± 18.73 |
233.9 ± 4.07 |
84.87 ± 6.466 |
27.8 ± 3.028 |
10.9 ± 1.067 |
Thr |
407 ± 30.14 |
239.9 ± 11.59 |
355.7 ± 2.423 |
58.36 ± 11.07 |
1.149 ± 0.366 |
Pro+β-Ala |
14.21 ± 0.639 |
28.73 ± 5.658 |
42.94 ± 8.26 |
17.98 ± 3.882 |
0.704 ± 0.231 |
Gly |
35.94 ± 2.387 |
26.26 ± 0.778 |
34.17 ± 2.539 |
40.21 ± 7.198 |
nd |
GABA |
105.8 ± 7.17 |
40.19 ± 1.837 |
84.76 ± 3.756 |
9.175 ± 1.978 |
7.235 ± 0.31 |
Ala |
253.4 ± 27.27 |
351.5 ± 5.481 |
387.7 ± 10.44 |
39.58 ± 6.501 |
2.866 ± 0.252 |
Val |
159.4 ± 9.839 |
155.4 ± 3.191 |
140.8 ± 4.633 |
50.55 ± 10.05 |
nd |
Met |
12.49 ± 0.862 |
4.772 ± 0.17 |
8.843 ± 0.528 |
0.461 ± 0.461 |
nd |
Leu |
37.07 ± 2.496 |
32.55 ± 2.118 |
25.58 ± 1.322 |
15.18 ± 2.881 |
0.16 ± 0.16 |
Ile |
45.71 ± 2.607 |
50.4 ± 0.83 |
57.76 ± 6.047 |
29.32 ± 5.613 |
nd |
Cys-Cys |
nd |
nd |
nd |
nd |
nd |
Trp |
3.819 ± 0.777 |
4.433 ± 0.697 |
nd |
11.58 ± 1.086 |
nd |
Orn |
2.935 ± 0.371 |
2.347 ± 0.009 |
nd |
3.12 ± 0.643 |
nd |
Phe |
26.63 ± 2.33 |
9.983 ± 0.101 |
15.8 ± 0.034 |
15.76 ± 2.987 |
nd |
Lys |
28.73 ± 3.061 |
17.24 ± 0.54 |
13.65 ± 2.164 |
9.518 ± 1.765 |
nd |
Cys |
1.588 ± 0.085 |
1.596 ± 0.055 |
1.26 ± 0.069 |
2.105 ± 0.142 |
nd |
Tyr |
49.61 ± 9.239 |
12.5 ± 0.283 |
9.671 ± 0.02 |
16.45 ± 3.433 |
3.353 ± 0.684 |
|
Cherry tomato |
Eggplant "Beinasu" |
Eggplant "Kamonasu" |
Eggplant "Kyokonasu" |
Eggplant "Marunasu" |
Eggplant "Zebranasu" |
His |
13.39 ± 5.62 |
31.46 ± 8.565 |
39.41 ± 3.139 |
88.56 ± 4.134 |
15.98 ± 2.025 |
30.67 ± 1.806 |
Arg |
19.27 ± 5.129 |
26.41 ± 1.75 |
80.53 ± 8.906 |
76.24 ± 2.669 |
16.53 ± 4.35 |
90.2 ± 8.215 |
HYP |
nd |
nd |
2.386 ± 0.125 |
nd |
nd |
nd |
Asn |
166.6 ± 71.7 |
355.4 ± 28.42 |
472.1 ± 63.92 |
992.8 ± 29.96 |
98.11 ± 19.69 |
152 ± 10.4 |
Gln |
472.9 ± 214.5 |
206 ± 22.98 |
269.1 ± 36.24 |
130.6 ± 7.851 |
89.43 ± 13.8 |
204.1 ± 10.87 |
Cit |
nd |
nd |
nd |
nd |
nd |
nd |
Ser |
48 ± 16.45 |
10.26 ± 0.677 |
25.77 ± 2.991 |
45.42 ± 2.354 |
10.78 ± 1.583 |
18.25 ± 0.891 |
Asp |
81.52 ± 41.2 |
29.51 ± 2.832 |
21.19 ± 0.597 |
60.67 ± 3.534 |
21.88 ± 2.575 |
25.66 ± 4.704 |
Glu |
687.4 ± 290.4 |
20.54 ± 1.197 |
20.41 ± 0.961 |
32.95 ± 2.064 |
17.63 ± 2.032 |
16.21 ± 1.467 |
Thr |
24.85 ± 7.489 |
23.64 ± 1.108 |
33.09 ± 3.141 |
62.48 ± 0.968 |
18.51 ± 2.113 |
20.87 ± 1.14 |
Pro+β-Ala |
22.29 ± 1.618 |
12.98 ± 0.407 |
17.22 ± 1.132 |
30.29 ± 1.987 |
9.24 ± 1.061 |
18.5 ± 2.429 |
Gly |
7.274 ± 2.767 |
4.403 ± 0.265 |
9.574 ± 0.55 |
7.413 ± 0.306 |
4.307 ± 0.463 |
7.901 ± 0.476 |
GABA |
93.59 ± 55.17 |
86.84 ± 7.584 |
221.5 ± 18.1 |
74.7 ± 3.888 |
184.2 ± 15.42 |
83.94 ± 6.821 |
Ala |
37.04 ± 16.65 |
3.619 ± 0.614 |
6.567 ± 0.482 |
9.47 ± 0.887 |
5.975 ± 0.901 |
8.224 ± 1.059 |
Val |
8.005 ± 3.296 |
21.67 ± 0.835 |
45.94 ± 2.538 |
61.48 ± 0.896 |
20.98 ± 2.597 |
35.03 ± 1.74 |
Met |
3.382 ± 1.15 |
nd |
nd |
nd |
nd |
nd |
Leu |
19.01 ± 6.342 |
4.738 ± 0.545 |
6.691 ± 0.134 |
10.81 ± 0.493 |
3.824 ± 0.9 |
16.46 ± 1.966 |
Ile |
7.223 ± 2.122 |
12.46 ± 0.639 |
20.22 ± 1.233 |
27.8 ± 0.593 |
11.12 ± 1.615 |
23.61 ± 1.435 |
Cys-Cys |
nd |
nd |
nd |
nd |
nd |
nd |
Trp |
1.438 ± 4.39 |
4.056 ± 0.645 |
7.87 ± 1.805 |
14.45 ± 1.306 |
2.713 ± 0.182 |
4.411 ± 0.285 |
Orn |
nd |
1.772 ± 0.13 |
7.206 ± 2.17 |
5.766 ± 0.577 |
1.46 ± 0.072 |
9.496 ± 1.016 |
Phe |
27.81 ± 6.499 |
17.34 ± 0.356 |
37.74 ± 1.647 |
97.05 ± 2.003 |
12.98 ± 1.54 |
18.1 ± 1.283 |
Lys |
8.565 ± 4.092 |
12.77 ± 1.369 |
10.92 ± 0.899 |
30.36 ± 1.895 |
8.691 ± 1.686 |
9.977 ± 1.494 |
Cys |
7.715 ± 0.878 |
nd |
nd |
1.626 ± 0 |
nd |
nd |
Tyr |
11.72 ± 2.1 |
8.541 ± 0.919 |
11.25 ± 1.104 |
31.58 ± 0.765 |
7.867 ± 1.1 |
9.702 ± 0.373 |
|
Spinach "Okame" |
Broad bean |
Cowpea |
Cowpea beans |
|
His |
nd |
126.6 ± 23.9 |
38.08 ± 6.123 |
163.6 ± 4.632 |
Arg |
0.372 ± 0.372 |
654.7 ± 54.74 |
38.2 ± 7.142 |
162.4 ± 3.91 |
HYP |
2.259 ± 1.311 |
nd |
nd |
nd |
Asn |
26.24 ± 1.131 |
945 ± 66.41 |
613 ± 30.48 |
392.6 ± 25.88 |
Gln |
35.31 ± 9.173 |
154.1 ± 4.755 |
551.4 ± 28.61 |
248.4 ± 8.96 |
Cit |
nd |
2.675 ± 1.693 |
nd |
nd |
Ser |
24.18 ± 1.985 |
142.1 ± 9.879 |
73.27 ± 2.145 |
72.32 ± 1.861 |
Asp |
48.59 ± 3.46 |
146.2 ± 13.99 |
46.51 ± 0 |
46.51 ± 0 |
Glu |
85.43 ± 7.502 |
181.6 ± 11.21 |
46.48 ± 1.804 |
265.7 ± 7.392 |
Thr |
9.409 ± 1.653 |
143.3 ± 14.71 |
45.86 ± 0.463 |
90.39 ± 1.414 |
Pro+β-Ala |
4.336 ± 0.561 |
36.5 ± 2.915 |
18.17 ± 0.047 |
67.29 ± 3.878 |
Gly |
1.165 ± 0.681 |
31.17 ± 1.757 |
6.967 ± 0.301 |
16.37 ± 0.785 |
GABA |
22.75 ± 1.147 |
226.6 ± 16.13 |
34.2 ± 1.668 |
15.39 ± 1.526 |
Ala |
21.94 ± 2.911 |
654.5 ± 64.95 |
31.8 ± 4.071 |
125.9 ± 3.136 |
Val |
4.754 ± 1.102 |
79.84 ± 2.213 |
41.82 ± 1.037 |
120 ± 4.047 |
Met |
nd |
1.7 ± 0.558 |
8.761 ± 0.393 |
25.52 ± 0.461 |
Leu |
3.405 ± 0.362 |
27.39 ± 0.715 |
21.44 ± 0 |
21.44 ± 0 |
Ile |
nd |
25.34 ± 0.754 |
23.29 ± 0.467 |
74.98 ± 2.901 |
Cys-Cys |
5.861 ± 3.487 |
nd |
nd |
nd |
Trp |
nd |
32.18 ± 2.075 |
3.813 ± 0.47 |
18.24 ± 1.499 |
Orn |
nd |
nd |
nd |
3.388 ± 0.124 |
Phe |
3.291 ± 0.61 |
11.6 ± 0.318 |
20.45 ± 0.577 |
58.47 ± 0.536 |
Lys |
nd |
60.26 ± 0.73 |
7.725 ± 0.576 |
17.8 ± 0.988 |
Cys |
109.3 ± 0 |
1.153 ± 0.126 |
nd |
2.286 ± 0.136 |
Tyr |
12.14 ± 1.411 |
60.24 ± 3.935 |
18.83 ± 1.493 |
72.12 ± 2.174 |
Amino acids are listed as abbreviated according to Table 1.
Average of 2~5 analyses (Means ± SE), nd: Not detected.
Table 5. Contents of free-form amino acids assayed in mushrooms (µmol/100 g sample on a wet weight basis)
|
Beech mushroom |
Beech mushroom
(Tanba) |
Button mushroom
(brown) |
Button mushroom
(white) |
Jew's ear |
King oyster
mushroom |
His |
29.38 ± 7.477 |
99.7 ± 5.005 |
43.63 ± 2.66 |
133.7 ± 3.481 |
9.663 ± 1.963 |
80.38 ± 3.689 |
Arg |
149.8 ± 8.816 |
311.2 ± 10.49 |
90.49 ± 2.751 |
99.8 ± 4.613 |
57.94 ± 13.16 |
302.6 ± 13.05 |
HYP |
2.095 ± 1.236 |
nd |
1.15 ± 0.734 |
1.847 ± 1.173 |
nd |
13.52 ± 0.789 |
Asn |
290.9 ± 31.47 |
113 ± 2.854 |
80.17 ± 3.27 |
178.1 ± 9.544 |
72.1 ± 12.47 |
106.5 ± 3.763 |
Gln |
149.8 ± 10.74 |
283.2 ± 17.75 |
322.7 ± 12.09 |
802.7 ± 41.74 |
68.81 ± 14.11 |
382.9 ± 15.61 |
Cit |
nd |
nd |
nd |
nd |
nd |
nd |
Ser |
60.76 ± 4.634 |
184.2 ± 7.715 |
101.5 ± 3.719 |
182.6 ± 8.61 |
14.09 ± 2.211 |
151.8 ± 3.506 |
Asp |
6.673 ± 0.615 |
7.738 ± 0.887 |
204.5 ± 8.97 |
210.3 ± 13.68 |
27.32 ± 6.626 |
37.37 ± 9.767 |
Glu |
222.9 ± 21.88 |
62.48 ± 7.219 |
299.3 ± 13.01 |
372.5 ± 29.14 |
26.04 ± 3.239 |
190.1 ± 16.83 |
Thr |
66.86 ± 4.54 |
121.2 ± 5.029 |
131.5 ± 5.477 |
206.8 ± 8.645 |
11.01 ± 1.4 |
145.7 ± 4.064 |
Pro+β-Ala |
24.26 ± 1.547 |
84.83 ± 2.042 |
97.13 ± 3.355 |
127.2 ± 4.381 |
2.392 ± 0.317 |
64.84 ± 1.564 |
Gly |
53.36 ± 3.844 |
141.4 ± 8.039 |
88.39 ± 3.428 |
163.2 ± 10.44 |
8.306 ± 1.602 |
129.2 ± 10.84 |
GABA |
nd |
98.47 ± 7.632 |
59.07 ± 2.68 |
140.9 ± 20.12 |
32.61 ± 8.053 |
35.12 ± 0.667 |
Ala |
86.72 ± 8.508 |
123.5 ± 9.437 |
240.8 ± 9.665 |
598.3 ± 50.35 |
43.95 ± 11.21 |
507.6 ± 61.07 |
Val |
33.9 ± 3.485 |
85.08 ± 5.763 |
94.7 ± 3.492 |
151.4 ± 8.963 |
4.24 ± 2.007 |
103.8 ± 6.616 |
Met |
nd |
15.67 ± 0.605 |
5.684 ± 0.26 |
8.567 ± 0.423 |
|
4.674 ± 0.199 |
Leu |
39.65 ± 4.621 |
96.53 ± 6.645 |
93.28 ± 3.86 |
144.5 ± 10.15 |
8.475 ± 3.134 |
144 ± 21.41 |
Ile |
18.91 ± 1.756 |
46.8 ± 2.982 |
46.45 ± 1.692 |
83.98 ± 5.49 |
3.667 ± 2.126 |
75.18 ± 6.733 |
Cys-Cys |
nd |
nd |
nd |
nd |
nd |
nd |
Trp |
nd |
nd |
nd |
32.86 ± 1.011 |
nd |
14.15 ± 0.772 |
Orn |
156.3 ± 20.96 |
242.2 ± 25.8 |
110.1 ± 8.37 |
98.03 ± 15.28 |
23.93 ± 8.903 |
199.2 ± 29.78 |
Phe |
38.54 ± 2.916 |
103.1 ± 3.677 |
100.2 ± 3.305 |
118.4 ± 4.251 |
nd |
73.4 ± 1.9 |
Lys |
12.8 ± 1.983 |
18.78 ± 3.628 |
25.74 ± 2.02 |
44.98 ± 7.77 |
6.433 ± 3.884 |
52.82 ± 9.439 |
Cys |
nd |
4.687 ± 0.72 |
nd |
|
nd |
1.773 ± 0.293 |
Tyr |
24.48 ± 1.605 |
75.12 ± 4.388 |
65.53 ± 2.636 |
50.22 ± 2.619 |
3.627 ± 0.616 |
76.28 ± 2.503 |
|
Mushroom "Honshimeji" |
Mushroom "Maitake" |
Mushroom "Shiitake" |
Porcini mushrooms (Dried) |
Winter mushroom |
|
His |
178.5 ± 10.14 |
41.05 ± 2.678 |
123.7 ± 5.593 |
527.7 ± 54.46 |
118.2 ± 3.371 |
|
Arg |
410.6 ± 18.82 |
47.35 ± 2.468 |
226.4 ± 12.59 |
724.1 ± 35.73 |
114 ± 3.03 |
|
HYP |
nd |
0.312 ± 0.312 |
nd |
244.2 ± 43.95 |
19.8 ± 1.552 |
|
Asn |
44.37 ± 3.853 |
16.44 ± 1.439 |
87.64 ± 3.504 |
1277 ± 142.8 |
101.7 ± 2.697 |
|
Gln |
755.9 ± 55.49 |
119.2 ± 9.674 |
1237 ± 83.49 |
629.6 ± 72.69 |
414.9 ± 11.71 |
|
Cit |
nd |
nd |
nd |
nd |
nd |
|
Ser |
93.53 ± 8.26 |
35.49 ± 3.123 |
97.06 ± 4.664 |
1499 ± 83.72 |
78.06 ± 2.904 |
|
Asp |
25.04 ± 0.899 |
11.71 ± 0.468 |
187.3 ± 15.24 |
325.5 ± 230.1 |
19.19 ± 3.483 |
|
Glu |
125.7 ± 4.428 |
75.59 ± 4.98 |
180.1 ± 13.53 |
462.6 ± 84.5 |
95.03 ± 3.556 |
|
Thr |
113.4 ± 8.527 |
42.18 ± 3.339 |
169.8 ± 10.55 |
694.9 ± 26.45 |
113.1 ± 4.342 |
|
Pro+β-Ala |
87.9 ± 6.65 |
17.79 ± 2.266 |
71.66 ± 1.074 |
396.4 ± 42.3 |
68.92 ± 3.138 |
|
Gly |
155.5 ± 6.678 |
21.9 ± 1.938 |
255.7 ± 15.22 |
1222 ± 144 |
123 ± 2.055 |
|
GABA |
46.12 ± 1.717 |
0.467 ± 0.467 |
80.73 ± 4.854 |
1378 ± 91.32 |
183.9 ± 5.589 |
|
Ala |
185.9 ± 6.48 |
92.2 ± 6.107 |
113.7 ± 6.708 |
2088 ± 148.1 |
252.6 ± 5.196 |
|
Val |
66.5 ± 3.585 |
21.51 ± 1.703 |
128.8 ± 7.157 |
262.2 ± 40.33 |
95.39 ± 1.62 |
|
Met |
10.53 ± 1.111 |
nd |
3.345 ± 1.156 |
346.9 ± 99.25 |
5.32 ± 0.205 |
|
Leu |
63.38 ± 3.125 |
8.217 ± 0.811 |
80.65 ± 4.762 |
65.07 ± 0.218 |
115.6 ± 5.381 |
|
Ile |
31.56 ± 1.551 |
11.55 ± 0.373 |
51.86 ± 2.891 |
92.72 ± 23.11 |
60.88 ± 1.216 |
|
Cys-Cys |
nd |
nd |
nd |
nd |
nd |
|
Trp |
nd |
nd |
nd |
103.6 ± 18.91 |
20.72 ± 0.598 |
|
Orn |
274.4 ± 13.8 |
62.71 ± 1.961 |
156 ± 14.37 |
66.86 ± 11.24 |
85.33 ± 2.289 |
|
Phe |
70.26 ± 4.225 |
22.54 ± 0.784 |
58.04 ± 2.473 |
123.4 ± 23.3 |
217.3 ± 5.183 |
|
Lys |
22.34 ± 1.081 |
7.209 ± 0.438 |
49.39 ± 5.58 |
96.48 ± 7.537 |
27.24 ± 1.79 |
|
Cys |
nd |
nd |
0.566 ± 0.19 |
8.504 ± 6.013 |
nd |
|
Tyr |
39.86 ± 1.959 |
24.42 ± 1.461 |
12.55 ± 2.442 |
136.9 ± 17.97 |
142.8 ± 6.058 |
|
Amino acids are listed as abbreviated according to Table 1.
The free-form amino acid compositions of five different kinds of eggplant were analyzed. Slight differences were observed in the free-form amino acid contents of the five species, but Asn, Gln, and GABA were abundant in all.
BCAAs, common ingredients in drinks designed for athletes, were not detected in most foods analyzed. However, moso bamboo and some peas, including the podded pea, contained considerable amounts of Val, Leu, and Ile BCAAs.
Here, we identified and measured significant levels of free-form amino acids in both vegetables and mushrooms. These free-form amino acids should contribute to the taste and palatability of foods. Indeed, Glu provides umami taste and has been shown to improve appetite [12,14,15], both of which contribute to QOL. Glu is particularly important in the foods of elderly and sick people, as it improves QOL by increasing nutrient intake. It is often discussed that excess intake of amino acids is toxic for human. For example, it has been warned that Glu has a potential risk of neurotoxicity like headache. However, this warning was almost eliminated after the joint report of FAO, WHO, and the Expert Committee on Food Additives (JECFA) in 2000 [28]. Thus, Glu is considered relatively safe and could be consumed without any concerns.
GABA is known for health-promoting functions, including an anti-hypotensive effect. Some studies suggest that supplementation with 10~20 mg (equivalent to 97~194 µmol) GABA reduces blood pressure in human [29,30]. Commercially available GABA enriched "Tokuho" foods are typically supplemented with 20 mg of GABA in Japan. Our present study suggests that two pieces of eggplant “kamonasu”, equivalent to 90 g, per day provides enough GABA to maintain blood pressure at a normal level.
Free-form amino acids are significantly beneficial to human health. The presence of these amino acids in vegetables and mushrooms suggests that they also provide a benefit to those species. In plants, free-form amino acids are likely absorbed directly from soil via the roots [31]. Some of these free-form amino acids are metabolized or stored, and some are utilized as signaling molecules to regulate cellular functions including enzyme activity, gene expression, and redox-homeostasis [32]. Pro, Arg, Met, Glu, and GABA are involved in the regulation of plant responses to various stress conditions [33-35]. Asn, the most abundant free-form amino acid in the foods analyzed in our study, is accumulated under stress conditions or mineral deficiencies [36]. In plants, free-form Asn also plays a central role in nitrogen storage and transport [36,37]. The roles of free-form amino acids in mushrooms are not well understood and are still being determined.
Here, we have demonstrated the presence of free-form amino acids in plants. However, there are many factors that influence the distribution of amino acids, including cultivation conditions, geographic production location, picking season, breed, and soil. In this study, we were unable to examine, or control, these variables. Nevertheless, we believe our results provide ample information with which to establish a free-form amino acid database to serve and inform nutritional studies.
- Kadowaki M, Torii K, Takahashi M (2013) Frontier of Amino Acid Science and Newly Applications for Better Human Life (Popular Edition), in Japanese, CMC Publishing, Osaka.
- Wu G (2009) Amino acids: metabolism, functions, and nutrition. Amino Acids 37: 1-17. [Crossref]
- Richard DM, Dawes MA, Mathias CW, Acheson A, Hill-Kapturczak N, Dougherty DM (2009) L-Tryptophan: Basic Metabolic Functions, Behavioral Research and Therapeutic Indications. Int J Tryptophan Res 2: 45-60.
- Fonnum F (1984) Glutamate: a neurotransmitter in mammalian brain. J Neurochem 42: 1-11. [Crossref]
- Legendre P (2001) The glycinergic inhibitory synapse. Cell Mol Life Sci 58: 760-793. [Crossref]
2021 Copyright OAT. All rights reserv
- McCormick DA (1989) GABA as an inhibitory neurotransmitter in human cerebral cortex. J Neurophysiol 62: 1018-1027.
- McIntosh TK, Vink R, Soares H, Hayes R, Simon R (1990) Effect of non-competitive blockade of N-methyl-D-aspartate receptors on the neurochemical sequelae of experimental brain injury. J Neurochem 55: 1170-1179.
- Roberts E, Frankel S (1950) gamma-Aminobutyric acid in brain: its formation from glutamic acid. J Biol Chem 187: 55-63. [Crossref]
- Crowe MJ, Weatherson JN, Bowden BF (2006) Effects of dietary leucine supplementation on exercise performance. Eur J Appl Physiol 97: 664-672. [Crossref]
- Diana M, Quilez J, Rafecas M (2014) Gamma-aminobutyric acid as a bioactive compound in foods: a review. J Funct Foods 10: 407-420.
- San Gabriel AM, Maekawa T, Uneyama H, Yoshie S, Torii K (2007) mGluR1 in the fundic glands of rat stomach. FEBS Lett 581: 1119-1123. [Crossref]
- Yamamoto S, Tomoe M, Toyama K, Kawai M, Uneyama H (2009) Can dietary supplementation of monosodium glutamate improve the health of the elderly? Am J Clin Nutr 90: 844S-849S.
- Tomoe M, Inoue Y, Sanbe A, Toyama K, Yamamoto S, et al. (2009) Clinical trial of glutamate for the improvement of nutrition and health in the elderly. Ann N Y Acad Sci 1170: 82-86. [Crossref]
- Toyama K, Tomoe M, Inoue Y, Sanbe A, Yamamoto S (2008) A possible application of monosodium glutamate to nutritional care for elderly people. Biol Pharm Bull 31: 1852-1854. [Crossref]
- Bellisle F, Monneuse MO, Chabert M, Larue-Achagiotis C, Lanteaume MT, et al. (1991) Monosodium glutamate as a palatability enhancer in the European diet. Physiol Behav 49: 869-873.
- Yamaguchi S, Ninomiya K (2000) Umami and food palatability. J Nutr 130: 921S-6S. [Crossref]
- Kato H, Rheu MR, Nishimura T (1989) Role of free aminoacids and peptides in food taste. In: Flavor chemistry: trends and development, ACS Symposium, Teranishi R, Buttery RG, Shahidi F (Eds.), ACS, Washington, D.C., pp: 158-174.
- Solms J (1969) The taste of amino acids, peptides and proteins. Int Z Vitaminforsch 39: 320-322. [Crossref]
- Kawai M, Sekine-Hayakawa Y, Okiyama A, Ninomiya Y (2012) Gustatory sensation of (L)- and (D)-amino acids in humans. Amino Acids 43: 2349-2358. [Crossref]
- Kagawa Y (2013) Standard Tables of Food Composition in Japan (in Japanese), Kagawa Education Institute of Nutrition, Tokyo.
- Moore S, Spackman DH, Stein WH (1958) Chromatography of amino acids on sulfonated polystyrene resins. An improved system. Anal Chem 30: 1185-1190.
- Spackman DH, Stein WH, Moore S (1958) Automatic recording apparatus for use in the chromatography of amino acids. Anal Chem 30: 1190-1206.
- Minamide T, Iwata T, Okino H (1985) The effect if stipe end cutting on freshness and composition of harvested nameko (pholiota nameko (T.Ito) S. Ito et. Imai) mushrooms (in Japanese). Nippon Shokuhin Kogyo Gakkaishi 32: 413-418.
- Sato E, Aoyagi Y, Sugihara T (1985) Contents of Free Amino Acids in Mushrooms (in Japanese). Nippon Shokuhin Kogyo Gakkaishi 32: 509-521.
- Suzuki T, Kubo N, Higinuma S, Tamura S, Yamamoto H (1981) Changes in free amino acid content during controlled atmosphere storage of lettuce, chinese cabbage and cucumber. In Rept Natl Food Res Inst, pp: 46-55.
- Suzuki T, Kurihara Y, Tamura S (1976) Amino acid content of fruits, vegetables and processed goods and similarity between their amino acid patterns. In Rept Natl Food Res Inst, pp: 42-70.
- Ito H, Ueno H (2014) Construction of free amino acids composition data base for food. J Chem Chem Eng 8: 501-515.
- Walker R, Lupien JR (2000) The safety evaluation of monosodium glutamate. J Nutr 130: 1049S-52S. [Crossref]
- Inoue K, Shirai T, Ochiai H, Kasao M, Hayakawa K, et al. (2003) Blood-pressure-lowering effect of a novel fermented milk containing gamma-aminobutyric acid (GABA) in mild hypertensives. Eur J Clin Nutr 57: 490-495.
- Shimada M, Hasegawa T, Nishimura C, Kan H, Kanno T, et al. (2009) Anti-hypertensive effect of gamma-aminobutyric acid (GABA)-rich Chlorella on high-normal blood pressure and borderline hypertension in placebo-controlled double blind study. Clin Exp Hypertens 31: 342-354.
- Näsholm T, Kielland K, Ganeteg U (2009) Uptake of organic nitrogen by plants. New Phytol 182: 31-48. [Crossref]
- Rai VK (2002) Role of Amino Acids in Plant Responses to Stresses. Biol Plant 45: 481-487.
- Ashrafa M, Harrisb PJC (2004) Potential biochemical indicators of salinity tolerance in plants. , 166: 3-16.
- Galili G, Höfgen R (2002) Metabolic engineering of amino acids and storage proteins in plants. Metab Eng 4: 3-11. [Crossref]
- Keutgen AJ, Pawelzik E (2008) Quality and nutritional value of strawberry fruit under long term salt stress. Food Chem 107: 1413-1420.
- Lea PJ, Sodek L, Parry MAJ, Shewry PR, Halford NG (2007) Asparagine in plants. Ann Appl Biol 150: 1-26.
- Postles J, Curtis TY, Powers SJ, Elmore JS, Mottram DS, et al. (2006) Changes in Free Amino Acid Concentration in Rye Grain in Response to Nitrogen and Sulfur Availability, and Expression Analysis of Genes Involved in Asparagine Metabolism. Front Plant Sci 7: 1-11.