In the interest of elucidating neural processes underlying memory, many have looked to computer information-theory and code, despite that these are inherently “demotive” i.e. lacking emotive qualities that render “meaning” or “value” to conscious experience. We review various textual coding schemes (i.e. Morse, Bar, QR) and consider their relevance to neural coding of “memory”.
We employ a psycho-chemical approach to describe the emotive neural memory code. The proposed tripartite mechanism and iconography involves the interactions of neurons with their surrounding extracellular matrix (nECM). Incoming perceptions are encoded into the nECM by neurons ejecting trace metal cations and neurotransmittters (NTs). They combine within local “addresses” in the nECM to form metal-centered complexes i.e. cognitive units of information (cuinfo). Neurotransmittter (NT) binding thereto confers emotive qualities to the cuinfo.
In short, the neuron employs chemo-affinity processes to encode (emotive) cognitive information. The nECM is the “library” while the metal cations and the NTs are the “dopants”. NTs are physiologic encoding elements, the molecular signifiers of emotive states.
Such a neuro-chemical code for memory has many credible features. Namely it:
• Employs the nECM surrounding neurons as a “library” of coding “addresses”.
• Employs dopants (metal cations and NTs) available to neurons, to encode units of memory.
• Encodes emotive (affective) states with NTs.
• Presents a multi-code (n>100), more complex than the binary code (n=2) of the computer.
• Entangles emotive memory with physiologic reactions.
• Is phylogenetically conservative. It starts with bacterial signaling and evolves with neural nets which ultimately achieves a phase change of “metabolic energy” into “mentality”.
We schematically represent the evolving chemo-affinity neural code as a phase transition of emerging mental talents of neural circuits.
coding, cognitive information, emotions, trace metals, neurotransmitters, memory
In the interest of elucidating the neural processes that underlie memory and learning, many have looked to the computer and its Information Theory. But there are pitfalls.
“It is probably dangerous to use this theory of information in fields for which it was not designed, but I think the danger will not keep people from using it” [1].
Here, we review various coding schemes and consider their relevance to neural signaling that results in memory. To begin, we define terms used to discuss both computer and neural memory.
Some aspire to describe neural memory and mentation in “objective” mathematical or algorithmic terms.
“Computational systems are useful to describe brain processes mathematically”.
[2].One could say that “data” is a quantifiable unit of knowledge, whereas “information” is vaguer. The single datum is the smallest unit of knowledge (input) which is physically represented in the chip (i.e. dopants distributed in a matrix [3,4]. By itself, an individual datum or a “bit”, is meaningless. But a set of bit data that integrates into a meaningful pattern is “information” (info). Thus, “information” derives from a material pattern encoding of “data”, which also requires the participation of a recognizing observer.
Equation 1.
Datum (singular) à Data (plural) à Pattern à Info (meaning)
recognition
For example, Hartley’s description [5,6] of the amount of information (info) in a message of length “L” using an alphabet with “s” symbols is:
Equation 2.
[Info] = sL
For a binary bit system where s=2, one could calculate:
Equation 3.
[Info] = 2L
But this approach divorces `’info” from” meaning”, as it is incapable of encoding the subjective import of stimuli. There is no binary calculus for emotions or “value” or “meaning”.
There have been many attempts to clarify how neurons encode memory. Many focused on an electrodynamic neural code subsumed under the terms “synaptic plasticity” , “long-term potentiation” and “action potentials (spikes)” [6-10]. Some attempted to combine electrodynamic processes with chemodynamic coding based on glutamate receptors (AMPARs) as underlying various forms of synaptic plasticity [11]. But these approaches did not entirely succeed in identifying the putative neural code.
It is the peculiarity of the neural system that sensations and memory are not described by mathematical formulae or by binary codes as in computers [4,12-16]. Some have argued that based on a computational model of the hippocampus,“top-down” expectation appear to interact with “bottom-up” input expectations [17].
But without memory, there can be no expectation. Thus, the argument is circular and the physiology of emotive memory remains vague. One could consider that the neuron employs a molecular code with neurotransmitters (NTs ) [18,19] to confer “meaning” or “value” to the emotive states elicited by sensations, as summarized in Table 1.
Table 1. Neurotransmitters (NTs), which elicit both physiologic reactions and psychic states
Neurotransmitter (NT) |
Physiologic reactions *
(perceptions, feelings) |
Emotive states |
Biogenic amines (8)
Amino acids (>10)
Neuropeptides (>70)
Acetylcholine (1)
NO (1)
Endocannabinoids (>10)
(trace metals; >10) |
Breathing
Blinking
Blood pressure
Blood coagulation
Cold (feel)
Contraction of muscles
Coughing
Cramps
Crying
Defecation
Dilation of muscles
Dilation of pupil
Drooling
Erection
Evacuation
Fever
Goose bumps
Heartbeat
Heat (feel)
Hunger (feel)
Immune reactions
Itching
Pain
Retching
Seeing
Shivering
Smelling
Thirst (feel)
Touching
Vomiting |
Anxiety
Aggression
Awareness
Craving
Curiosity
Depression
Desire
Dread
Dreams
Fantasy
Fear
Hate
Joy
Love
Paranoia
Sadness
Sex drive
Sociability |
|
* No memory required. |
! Emotions requiring memory |
New Dimension
One could consider that the neural net instigates a new dimension of biologic energy, a mental state of psychic proportions, recalled as memory. The transformation of sensation to cognitive information (cog-info) could be formulated as:
Equation 4.
Sensum àSensation à Emotive qualityà Cog-info
NTs
where analogous to Equ.3 of info, one can formulate cog-info as:
Equation 5.
[Cog-info] = 100 L
The emotive content of stimuli is a key difference between the binary info of the computer chip and the “multinary” cog-info of the neuron. In that the messages (perceptions) experienced by neural nets are not discrete but continuous, the term L used above (equations 3 and 5) is somewhat pointless. But it reveals the complexity of the neural experience. Bottom line.
Equation 6.
[Info] ≠ [Cog-info]
[Cog-info]
Indeed, trying to rationalize the psychic dimensions of neural experience with text or mathematical algorithms is reminiscent of 2D shapes (circles) trying to comprehend a 3D reality (spheres), as parodied in “Flatland” more than 100 years ago [20]. On a more theologic tack, unwillingness to rationalize mental states as physiologic events may incline homo religiosus to cast mentality in terms of “Spirit” couched in Talmudic dielectics [21-23]. It may also underlie the paradoxical position of chemically-minded scientists who describe reproduction and life processes as involving transition metal complexes, but who hesitate to ascribe the emergence of
mental states to similar biochemical reactions [26,27].
Indubitably, neural processes are emotive with affective, as well as objective, qualities. For example, pain is perceived with an attendant psychic state that is remembered to avoid (See conditioning a la Pavlov, Skinner). To date, no-one has written a mathematical formulation for pain, love, fear, etc. Artificial intelligence (AI) [28-31] provides no codes, formulae or algorithms that simulate an emotive state experienced by a neural creature; for humans, these are expressed in poems, plays, music and prayers. Rather than look to the Information Theory derived from Babbage, Boole, Turing, von Neumann, Shannon, Wiener and their like [29,30], one must employ physiologically relevant descriptions of an evolved, chemo-affinity neural code [31,32] aware that like a Russian doll, mentation by a brain operates at various scales of biological organization (anatomic, cellular, membranal), but with all compartments ultimately operating on a biochemical (molecular) scale.
Like the computer which encodes “info” as physical bits, the neural net too encodes “cog-info” physically dare one say, chemically. Here, we review various coding systems and consider how they might apply to neural net signaling which results in memory.
The term “code” has been applied in jurisprudence to refer to to laws that define acceptable social behavior (i.e. Napoleonic code). For signaling purposes, other meanings apply i.e:
- A code involves replacing one letter or word or phrase for another.
- A process of instructions for treating binary, octal, or hexadecial formatted data to be deciphered and executed by a computer, subsequently translated into text [33-35].
Alphanumeric code: employed to relay secret text messages or to encode passwords. Examples are the famous Caesar code, which was based on the rote displacement substitution of letters. More complicated codes have been devised (see the German Enigma machine of WWII, whose code was broken by Marian Rejewski (a Pole) and Alan Turing (an Englishman)). But they are essentially linguistic manipulations (Equation 7) to decipher a cryptic textual code into plain meaning text (Keyence manual) and do not relate to neural signaling.
Equation 7. “Enigma” cryptography process:
text à TEXT à text cracked
plain cryptic plain
The morse code: (Figure 1) Originally developed for the telegraph to convert electrical signals to text, as a binary code (dots, dash), is formatted with 5 characters separated by a space (pentad code). The dot-dash-space coding convention permits representations of 36 letters and numbers.

Figure 1. Morse code translates dots and dashes into alpha-numeric text
Equation 8.
Morse binary (pentad) coding options = b1 + b2 + b3 + b4 + b5 = 62
where b = 2 (dot/dash or binary 0 1).
Any code that employs more than 2 effectors (i.e. dot, dash) can be considered as multiplexed. For example, if one were to employ a multitude of effectors (i.e. b=10) to encode a 5-digit message of Equation 8, one would obtain many text coding options
Equation 9.
Multinary (pentad) coding options = b1 + b2 + b3 + b4 + b5 = 111,110
where b = 10.
Clearly, increasing the number of coding effectors for text messages exponentially increases the encoding options.
Bar code: A liner bar code (1D) contains only about 30 characters to express product information. This bar code is machine readable and widely used for identifying inventory, tickets and luggage. But is “demotive” in that it cannot encode an emotive state (Figure 2).

Figure 2. Bar code, read by laser scanners
QR code: In response to the need for greater coding capacity, the 2D QR matrix code was developed, which has up to a hundred times the data capacity, even though it is over ten times smaller [36-38]. The more complex QR codes were initially developed by Japanese industry and later adapted to a large range of uses. For many applications, QR icons (Figure 3), which are read by laser scanners, encode in 4 modes (numeric, alphanumeric, binary and kanji), comprising small squares arranged in a square grid.

Figure 3. QR code has a square frame, for reliable position detection and more efficient data storage than the simpler bar-code; its speed is its most important quality. These machine codes are totally “demotive”, incapable of encoding emotive states
DNA/RNA triplet codon code
Involves a slow read/write process that requires high energy and a complex molecular translation process. The quadro-triplet genetic information of DNA (i.e. triplets of 4 bases) can be transferred or even synthesized in vitro and is not necessarily lost by the host’s death (practiced as ‘molecular biology’ [24]. While it can code for the 64 amino acids for the biosynthesis of proteins or for small sequences to perform as switches in biologic processes, it is limited in that it cannot signify the complex emotive quality of neural memory.
Immunologic code
The recognition of antigens by antibodies and cell sensors of such are the basis of immunologic monitoring of the body. The antigen/antibody system can be considered to “remember “ previous insults; there are specialized lymphocytes that remember these [37]. Though it can instigate pain, discomfort and organ rejection, the antigen/antibody code does not translate into mentality or conscious memory.
Neural Codes/Signals
By contrast to synthetic codes, neural coding/signaling is not at all linguistic, but “experiential”. Thus, for neurons interacting within a circuit, one must expand the meaning of “code” to include “signals” with whole-body reactions associated with mental states (Table 1). Many workers assume that the neural code refers to the timing or magnitude of electrodynamic signals (spikes) [34,38,39].
It may be that attempts to describe emotive states analytically/linguistically are doomed to fail, nalogous to Kurt Godel’s demonstration that even some mathematical statements are beyond the reach of logical descriptions [3,36,40,41]. Stoo, emotive states are beyond the reach of logical descriptions. To be applicable to all neural creatures that exhibit the talents of emotions and recall [42], one needs to describe a biochemically credible mechanism and identify relevant signalling effectors.
While most attention has been directed at synaptic electrodynamic signaling between neurons [43-47], non-synaptic chemical signaling cannot be ignored but must be accounted for in any complete description of neural signaling [32,48,49]. Numerous efforts have been expended on trying to conceptualize the code that neurons employ to effect memory, as summarized in a sampling below:
Signaling pathways: It has been suggested that “information for learned behavior of biological systems may be stored within intracellular biochemical reactions that comprise signaling pathways.” [50].
While they discuss rate and equilibria constants, as well as signaling loops and reversible/ irreversible reactions, they remain vague on the persistence of memory and the coding details, particularly how emotive states are encoded.
Long-Term Potentiation (LTP) has been forwarded as a process underlying the synaptic plasticity (SP) that many contemporary neuroscientists believe is a core component in the mechanism of memory. Its roots lie in the work of Cajal and later Hebb (1911 and 1949 respectively) [5,53]. But such analysis is incomplete, requiring further consideration of the meaning of neural morphology and immediate surroundings, the kinetics of morphologic changes related to learning and how they impact on the plasticity of synaptic functionality.
Morphogenesis and synaptic connections say little about how neural nets evolved to achieve psychic states. One could consider that neural signaling evolved from bacterial signaling and still uses the same molecules i.e. biomodulators now called neuro transmitters (Table 1). A recent review summarized the difficulties of ascribing cognitive function, such as emotive memory, to electrodynamically connected sets of neurons [52].
Sugar code
It has been recognized that poly-sugars, as glycan-based interactions might serve as a coding system which exquisitely regulates basal nerve processes such as axon growth, neuron, differentiation and synaptic signaling [55,56]. Most work focused on biosynthesis, development and morphogenesis, but little on a neural signaling code.
Sulfation patterns of heparan sulfate (HS). Some [55] seek to understand how cell type and development are related to specific HS sulfation patterns. For neurons, it seems apparent that the degree and distribution of sulfate groups in GAG could affect transition metal cation binding. But its relevance to neural encoding of memory remain to be explored.
A quantum physics approach has been suggested as having a direct impact on, or being directly responsible for, cognitive function [55-57]. However, quantal effects are “demotive” and it is hard to consider them at a physiologic level. Though both quantum mechanics and cognitive psychology are awash in uncertainties and entanglements, they are not causally related phenomena and do not explain one another [58].
The pioneering ideas of mathematical biologists [58-62], notably Alfred Lotka [61].
“A state of consciousness (i.e. memory) can be described either in terms of its “contents” or in terms of the disposition of the molecules etc of the brain, just as a magnetic field might be described either in terms of an intensity chart or the position of a number of magnets.
“Consciousness has evolved… spun into a continuous thread by a faculty of memory”.
One cannot really object to these phrases which were written a generation ago. But they are inadequate to a modern neuroscientist looking for a physiologically relevant causal mechanism that results in neural memory rooted in evolved biology.
Axiology is a direct, multi-faceted description of a structure and function of the human language. It has been suggested that it encapsulates a “G code” which defines how the brain works to generate language [26,60].
Comment
Though language might be considered as a code to remember and to emote, it applies only to humans. Linguistic analysis [61] does not clarify the universal, physiologic mechanism underlying neural memory or reveal its phyto-evolutionary roots. Psycho-linguistic analysis, namely the retrieving, parsing and processing of a patient’s discourse to generate an “axiological” value of a patient’s mood state [60] does not clarify how a mood state is encoded or remembered.
The neural electrodynamic signaling code has the form of structured net fragments that are formed by network self-organization i.e. “excitory connections between neurons”. This neural network concept raised two important questions:
- What comprises the network, whose “structural fragments” are essential for
the elicitation of a state of mind?
- What is the mechanism through which experience forms memory?
The questions were valid but remained unanswered. Thus, graphic representations of 2-dimensional sheets of dots connected by lines or as tracings of spiking patterns do not satisfy the need for a physiologically relevant accounting of neural signaling which results in mentality. Moreover, these approaches cannot identify the
means by which emotive states are encoded or recalled [62-64]. Similarly, the problem of producing man-made "machine" consciousness and "artificial thought” including memory, was considered as a matter of duplicating
all levels of the operational architectonics of brain electromagnetic signaling [65-67]. Though purported to be “neurophysiological”, it focused entirely on the electrodynamic activities of the brain, and ignored all the chemical signaling performed by active neurons. Neither did it address the issue of “emotive states” experienced by biological neural nets.
In hypothesizing how groups of neurons process sensorial information, some neuroscientists have adopted questionable terms and concepts . For example, neurons do not “compute”.They do not process information (info) in binary terms (01). Rather, they “mentate” cognitive information (cog-info) with emotive qualities (discussed below).
With regard to “population coding”, a “thesaurus” of a neural population code that serves to encode information by eliminating neural network noise, (44, was suggested Ganmor et al, 2015; Averbeck et al, 2015). This is incomprehensible due to a misuse of words and assumptions regarding the nature of neural signaling, which is not verbal. Thus, employing words like “semantic,” “thesaurus” or “vocabulary” is meaningless in the context of neural signling [68]. A “correlation” of the signalling activity of a group of neurons (i.e. an anatomic compartment) in response to a stimulus does not in itself serve to decode the signals resulting in a mental state.
For example, in the Atlantic battles of WWII, simply “detecting” communications between submarines and their German headquarters was in itself not a “decoding”, though it “correlated” with the submarine attack activity (Figure 4).
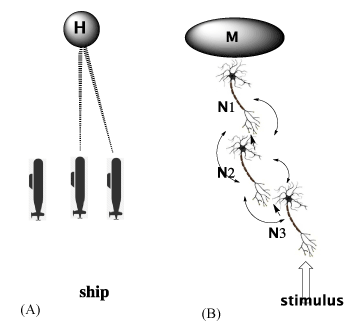
Figure 4A. Schematic of 3 submarines (S1, S2, S3) signaling encoded text to each other and to headquarters (H) during WWII. Note that the presence of surrounding water is ignored and left blank. Detection of the frequency of their encoded signals may have correlated with attacks on ships, but did not mean that the underlying messages were deciphered. Rather, it required a cracking of the Enigma code (text à TEXT à text) to comprehend the signaled messages
Figure 4B. Schematic of 3 neurons signalling one another. Note that the surrounding nECM, through which non-synaptic signalling (curved arrows), is not represented (left blank), but ephaptic signalling through the nECM represented by curved arrows
Similarly, correlating the frequency of synaptic signalling to stimulus does not clarify how a psychic state of mind or memory (M) is achieved.
Proposed: Tripartite mechanism of encoding neural memory
In an attempt to specify the “nuts and bolts” of neural memory in atomic units and scale, we have proposed a tripartite mechanism that involves the interactions of neurons with their surrounding extracellular matrix (nECM) [71-73]. Thus, neurons are not “naked”, but enshrouded in a web of glycoaminoglycans which serve as its “memory material”. Incoming perceptions are encoded with trace metals + neurotransmitters (NTs) Table 1 to form metal-centered cognitive units of information (cuinfo). In conjunction with whole body effects, the NTs are considered to be the effectors and signifiers of emotive states achieved by stimulated neural nets. Alternatively phrased, the NTs are the coding “alphabet” of emotive states and the physiologic reactions are the “syntax” [65]. Table 1 summarizes the entangled physiologic and psychic effects of NTs [69-71].
The chemographic notation in Figure 5 [71-73], encapsulates the general coding features of the tripartite mechanism. It remains cryptic in that it does not specify how specific NTs elicit various emotions. But it identifies the salient components of neural signaling system that entangles psychic states with physiologic reactions.
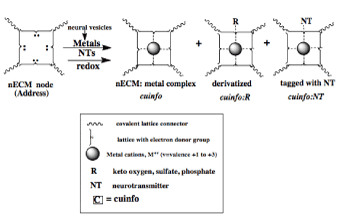
Figure 5. Chemographic representations of the reaction of a nECM binding site for a metal cation, an electron rich site (“address”}. The binding of a neurotransmitter (NT) to the metal-centered cognitive unit of information (cuinfo) confers emotive context
Thus, the tripartite mechanism, which describes a chemical “code” for psychic states, is not accessible to synthetic devices or simulations. The cuinfo are the molecular correlates of a memory, units (also referred to as engrams) that render the synaptic connections operative [72]. Neural signaling is based on molecular interactions [31,73] instigated by neurons interacting with their surrounding nECM. Essentially, the entire “edifice” of memory rests on the proper functioning of a chemo-affinity signaling process [28] involving metal-centered complexes within the nECM, the neuron’s “memory material”.
The metaphor of the computer for the brain is flawed. Artificial intelligence researchers have eriodically considered adding emotions to computers and robots so as to prioritize and direct their processing [28-31]. But faced with inability to describe emotive cognitive information with a binary code or a mathematical algorithm, this approach has come to naught [41]. According to the laws of quantum mechanics, information may never be destroyed. But the cognitive information of most neural creatures disappears when they die. It is not inherited or genetically transmitted to their offspring via the DNA “code”. The special case of humans relates to the inheritance of some cognitive information in the form of literature, poems, music paintings and media (movies, YouTube etc.). Some have suggested that the capability to integrate information is the basis for neural consciousness [74,75]. We point out that the “sense” or “meaning” of computer data is determined by the programmer who establishes the way in which data organized into valued information.
Q: How is a pattern recognized as information? Who does the valuation?
Another aspect that needs to be considered is that decrypting the “neural code” (if it can be so called) is not like decrypting linguistic codes/ciphers (i.e. monoalphabetic substitution cipher, or the homophonic substitution cipher, or the polyalphabetic cipher, or the Vigenere cipher, or the Enigma code [33]. All these essentially translate known text into hidden text, or vice versa. However, the neural signaling code is not linguistic but physiologic and biochemical. It relates to signals which elicit (emotive) mental states of being, for which there are no words or textual equivalents, only manifest as other dimensions of experience.
The cuinfo (Figure 5) are the “chemo-affinity” inscriptions of perceptions . Even so, identifying the chemical “script” does not mean that one has “cracked” the “neural code”. That is, even graphic representations cannot penetrate subjective, psychic experience. Like the physicists’ inability to approach a black hole closer than its “event horizen” [76,77], the neuroscientist may not be able to render a rational account of the mental states achieved by neural nets, which are beyond words, numbers and graphs, but not of drugs (i.e. Prozak, cocaine, etc).
Consider the meaning of “meaning”. From where does ”meaning” of a computer message derive? In that the bit information message is inherently “demotive”, it cannot convey an emotional state or valuation. Rather, the value of a message arises only in the brain of the recipient neural creature, whose very existence depends on its ability to render judgement as to message “worth” or “meaning”, ultimately based on emotive valuation.
The gap between “information” and “knowledge” is wide. The former lies in the realm of computers and mathematics; the latter can only be achieved by neural creatures driven make value judgements to survive the “experiential phase” of life and to reproduce. But both activate a recall function called “memory”, considered to be “the intersection of mind and matter” [78]. But details are lacking and open to philosophic conjectures (as by Bergson or Husserl, in [80,81]). It has been posited that “information” recognized by “mind” is necessarily physical, as a pit, hole, spin, dopant etc. Analogously, we propose that “cognitive information” can be chemographically represented chemically, as in Figure 5.
Mind from matter
Hopefully, we avoid Ryle’s “category error” (a logical fallacy that occurs when one makes an argument against Mind/Body dualism [80-82] by assigning a physical (chemical) process to an immaterial talent, namely neural memory. The neuro-chemical code we propose has many credible features lacking in others, namely:
- Provides a multi-code (n>100) distinct from the binary code (n=2) of the computer.
- Identifies a means for encoding emotive states with NTs.
- Employs NTs to entangle physiologic reactions with psychic states.
The emergence of memory and the evolution of neural systems with higher cognitive talents from simpler cells can be considered as a phase transition, with the emergence of mental talents related to the number of neurons in the brain [83-86], as schematically represented in Figure 6.
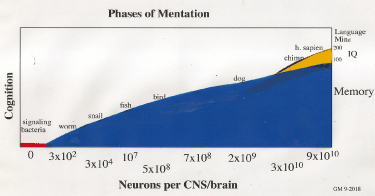
Figure 6. Phase changes in the evolution of memory from bacterial signaling to higher cognitive functions in h. sapien. The red region  describes bacterial signaling with modulating molecules, notably biogenic amines, amino acids, acetyl choline and NO (Table 1); blue region
describes bacterial signaling with modulating molecules, notably biogenic amines, amino acids, acetyl choline and NO (Table 1); blue region  relates to neural memory involving signaling with additional neuropeptides (Table 1); orange
relates to neural memory involving signaling with additional neuropeptides (Table 1); orange  refers to higher cognitive talents emerging from more complex neural signaling. Of course, trace metals and NTs are available to all these biologic systems [87,88]
refers to higher cognitive talents emerging from more complex neural signaling. Of course, trace metals and NTs are available to all these biologic systems [87,88]
Heuristic Implications of the tripartite mechanism
Where do we go from here? It has been pointed out that the value of a new scientific thesis is that it suggests new experimental approaches to the subject under consideration. For us, the subject is the mental talent of neural memory, for which we have proposed a tripartite mechanism. Thus, we have undertaken to experimentally test some aspects of this mechanism by evaluating the effects of coating impedence electrodes with components suggested by the tripartite mechanism, namely neurotransmitters (NTs) and glycosaminoglycan (GAG) nalogues of the nECM components that surround neurons.
Results with oxytocin-coated electrodes were positive for detecting low levels of trace metals, as recently reported (Figure 7) [85]. Electrodes were also coated with sulfated tetra-saccharide analogues [87,88] of hyaluronate, exhibited unique sensitivity to different race metal cations [86].

Figure 7. Schematic of a coated impedence electrode that can selectively detect low levels (10-12 M) of trace metals Cu+2 and Zn+2. The electrodes was covalently-coated with oxytocin (OT) or sulfated saccharide. The resultant metal- complexes could modulate the electrode responses
The brain performs much better than a computer in many respects. It may be slower at arithmetic processing but it comprehends existential circumstances far beyond the ken of the computer. The lack of emotive coding effectors prevents the computer from encoding emotive algorithms. It can only encode with 0 or 1, with no emotive significance. Recognizing a face or a voice does not make the computer affective [30]. It may have been programmed to identify emotive states in man, but that is not equivalent to an emotive experience. It is apparent that neural sensibility which is inherently emotive, addresses a dimension not accessible to computers. Possibly, such a possibility will expand the computer horizens beyond the limitations of Moore’s Law [87]. Such experimental techniques go a long way to buttress the tripartite concept and lend credibility to this biochemical approach to a heretofore unapproachable mental process [86-88].
I thank my nephew JJ Marx Esq (Jerusalem) for suggesting that we include ideas for future experimental directions. We note that Professor Gallistel’s remarks (Rutgers University) drew our attention to “memory” as the proper focus of our speculations. Thanks to my companion Karine Ahouva Leopold (Jerusalem, Paris) for introductions, emotional support and encouragement. We appreciate Professor Jean Paul Doguet’s (Paris) interest and suggestions regarding the relevance of the philosophers Husserl and Bergson. We also thank Prof. Poznanski (Universiti Sultan Zainal Abidin, Malaysia) for drawing our attention the works of Lotka.
GM is a founder of MX Biotech Ltd., with the commercial goal to develop new “memory materials” and devices. CG is emeritus professor at the Institute of Chemistry, The Hebrew University of Jerusalem. He is active in developing technologies for the conversion of peptides and active regions of proteins into orally available drug leads is active in developing and patenting peptide-based tools for surgery and pharmacology. Notwithstanding, the ideas forwarded here are scientifically genuine and presented in good faith, without commercial clouding of the concepts expressed here.
- Licklider JCR (1950) In: Claude E. Shannon "The Redundancy of English". In: Pias, C; von Foerster, H. eds. 2003. Cybernetics: Transactions.Published by Diaphanes: Zürich, Berlin.
- Guidolin D, Albertin G, Guescini M, Fuxe K, Agnati LF (2011) Central nervous system and computation. Q Rev Biol 86: 265-285.
- Turing A (1937) On computable numbers with an application to the Entscheidungs problem. Proc London Math Soc 42: 230-265.
- Hartley RVL (1928) How the rate of transmission of information over a system is limited by the distortion resulting from storage. Bell System Technical Journal7: 535-563.
- Hebb DO (1949) The Organization of Behavior Wiley, New York.
- Azarfar A, Calcini N, Huang C, Celikel FZT (2018) Neural coding: A single neuron’s perspective. Neuroscience & Biobehavioral Reviews 94: 238-247.
- Freeman WJ (2010) The use of codes to connect mental and material aspects of brain function comment on: “Natural world physical, brain operational, and mind phenomenal space-time” by A.A. Fingelkurts, A.A. Fingelkurts and C.F.H. Neves Physics of Life Reviews 7: 260-261.
- Panzeri S, Harvey CD, Piasini E, Latham PE, Fellin T (2017) Cracking the neural code for sensory perception by combining statistics, intervention, and behavior. Neuron 93: 491- 528. [Crossref]
- Sardi S, Vardi R, Sheinin A, Goldental A, Kanter I (2017) New types of experiments reveal that a neuron functions as multiple independent threshold units. Sci Rep 7: 18036.
- Diering GH, Huganir RL (2018) The AMPA receptor code of synaptic plasticity. Neuron 100: 314-329. [Crossref]
- McCulloch WS, Pitts W (1943) A logical calculus of the ideas immanent in nervous activity. Bulletin of Mathematical Biophysics 7: 115-133.
- Reprinted (1988) In: McCulloch, Embodiments of Mind. MIT Press, Cambridge, MA.
- Marr D (1971) Simple memory: A theory for archicortex. Philos Tansact Royal Soc London B Biol Sci 262: 23-81. [Crossref]
- Fodor JA (1975) The language of thought. Harvard University Press, Boston, MA.
- Gallistel CR, King AP (2009) Memory and the computational brain. Wiley Blackwell, New York.
- Street S (2016) Neurobiology as information physics. Front System Neurobiol 10: 90.
- Frank D, Montemurro MA, Montaldi D (2020) Pattern separation underpins expectation-modulated memory. J Neuroscience 40: 3455-3464.
- Dulcis D, Jamshidi, Leutgeb S, Spitzer NC (2013) Neurotransmitter switching in the adult brain regulates behavior. Science 340: 449-453.
- Werner G, Mitterauer BJ (2013) Neuromodulatory systems. Front Neural Circuits 7: 1-4.
- Abbott EA (1884) Flatland: A Romance of Many Dimensions. Seely & Co. Ltd, London, Reissued as Dover Publication, 1992, New York.
- Spinoza B (1994) A Spinoza Reader: The Ethics and Other Works 1677. Edited and translated by Curley E., Princeton University Press, Princeton, NJ.
- Lin M (2005) Memory and personal identity in Spinoza. Canadian J Philos 243: 243-268.
- Soloveitchik JB (2005) Halakhik Man. English translation from Hebrew, L Kaplan. Sefer ve Sefel Publishing, Jerusalem.
- Watson J, Baker TA, Bell SA, Gann A, Levine M (2013) Molecular biology of the gene. 7th Edition. Pearson PublishersNew York. ISBN-13: 978-0321762436.
- Chang CJ (2017) Bioinorganic life and neural activity: Toward a chemistry of consciousness? Acc Chem Res 50: 535-538. [Crossref]
- Crick F, Koch C (2003) A framework for consciousness. Nature Neuroscience 6: 119-126.
- Franklin S (1995) Artificial Minds. MIT PressCambridge, MA.
- Howard N, Guidere M (2012) LXIO: Mood Detection Robopsych. Brain Sciences Journal 1: 69-75.
- Bosse T, Broekens J, Dia J, van der Zwaan J (Eds.) (2014) Emotion Modeling. Towards Pragmatic Computational Models of Affective Processes. Springer, New York.
- Hudlicka E., From habits to standards: Towards systematic design of emotion models and affective architecture. pp 3-23.
- Dastani, M.; Floor, C.; Meyer, J.J. Programming agents with emotions. pp 57-75.
- Lowe R.; Kiryazov, K. Utilizing emotions in autonomous robots: An inactive approach. pp 76-98.
- Sperry RW (1963) Chemo-affinity in the orderly growth of nerve fiber patterns and connections. Proc. Natl. Acad. Sci. USA 50: 703-711.
- Hasson C (2011) Modelisation des mecanismes emotionnels pour un robot autonome: Perspective developpementale et sociale. Universite de Cergy Pontoise, PhD Thesis, France.
- Robinson P, Kaliouby R (2009) Computation of emotions in man and machines. Philos Trans Roy Soc Lond B Biol Sci 364: 3441-3447.
- Jaross W (2015) Hypothesis on a signalling system based on molecular vibrations of structure forming macromolecules in cells and tissues. Integr Mol Med 2: 361-364.
- Jefferys JG (1995) Nonsynaptic modulation of neuronal activity in the brain: electric currents and extracellular ions. Physiol Rev 75: 689-723.
- Singh S (2000) The Code Book: The secret history of codes and code-breaking. Fourth Estate, London.
- Softky WR (1995) Simple codes versus efficient codes. Curr Opin Neurobiol 5: 239-247.
- Shiffman D (2012) The nature of code. Magic book project, New York.
- QR ID Code Handbook (2019) Keyence manual. Basic practice of 2D codes. Keyence corporation of america publisher.
- Ratajczak W, Niedźwiedzka-Rystwej P, Tokarz-Deptuła B, Deptuła W (2018) Immunological memory cells. Cent Eur J Immunol 43: 194-203.
- Shadlen MN, Newsome WT (1994) Noise, neural codes and cortical organization. Curr Opin Neurobiol 4: 569-579.
- Urgolites ZJ, Hopkins RO, Squire LR (2019) Spared perception of the structure of scenes after hippocampal damage. J Cogn Neurosci 31: 1260-1269.
- Godel K (1932) On formally undecidable propositions. Basic books, New York (translation of 1932 paper).
- Pockett S (2014) Problems with theories that equate consciousness with information or information processing. Front Syst Neurosci 8: 225.
- LeDoux JE (2000) Emotion circuits in the brain. Annu Rev Neurosci 23: 155-84.
- Asok A, Leroy F, Rayman JB, Kandel ER (2019) Molecular mechanisms of the memory trace. Trends in Neurosciences 42: 14-22.
- Ganmor E, Segev R, Schneidman E (2015) A thesaurus for a neural population code. ELife 4: e06134.
- Costandi M (2017) Neuroplasticity. MIT Press, MA.
- Li M, Tsien JT (2017) Neural Code-Neural self-information theory on how cell-assembly code rises from spike time and neuronal variability. Front Cell Neurosci 11: 236.
- Kandel E, Dudai Y, Mayford MR (2014) The molecular and systems biology of memory. Cell 157: 163-186.
- Bokil H, Laaris N, Blinder K, Ennis M, Keller A (2001) Ephaptic interactions in the mammalian olfactory system. J Neurosci 21: RC173.
- Anastassiou CA, Perin R, Markram H, Koch C (2011) Ephaptic coupling of cortical neurons. Nat Neurosci 14: 217-23.
- Bhalla US, Iyengar R (1999) Emergent properties of networks of biological signaling pathways. Science 283: 381-389.
- Cajal RY (1911) Histology of the nervous system of man and vertebrates. Oxford University Press,1995 ISBN: 9780195074017
- Arshavsky YI (2017) Neurons versus Networks: The interplay between individual neurons and neural networks in cognitive functions. Neuroscientist 23: 341-355.
- Gabius HJ (ed.) (2009) The Sugar Code. Fundamentals of Glycosciences. Wiley, New York.
- Abad-Rodriguez J, Diez-Revuelta N (2015) Axon glycoprotein routing in nerve polarity, function and repair. Trends Biochem Sci 40: 385-396.
- Lamanna WC, Kalus I, Padva M, Baldwin RJ, Merry C (2007) The heparanome—The enigma of encoding and decoding heparan sulfate sulfation. J Biotechnology 129: 290-307.
- Penrose R (1987) Quantum physics and conscious thought in Quantum implications: Essays in honour of David Bohm (Ed. B.J. Hiley and F.DE. Pearl). Routeliedge & Kegan Paul, London.
- Penrose R (1989) The Emperor’s new mind. Oxford university press. London, UK.
- Poznanski PR, Cacha LA, Latif AZ, Salleh SH, Ali J (2019) Theorizing how the brain encodes consciousness based on negentropic entanglement. J Integrative Neuroscience 18: 1-10.
- Lotka AJ (1925) Elements of physical biology. Williams and Wilkins Company London.
- Howard N, Hussain A (2018) The fundamental code unit (FCU) of the brain: Towards a new model for cognitive geometry. Cognitive Computation 10: 426-436.
- Chomsky N (1975) Reflections on Language: Harper Collins Distribution Services, UK.
- Mejia JF, Longtin A (2012) Optimal heterogeneity for coding in spiking neural networks. Phys Rev Lett 108: 228102.
- Dahaene S (2014) Consciousness and the Brain. Deciphering how the brain codes our thoughts. Viking Press, New York.
- von der Malsburg C (2018) Concerning the Neuronal Code. J Cognitive Science 4: 511-550.
- Freeman WJ (1997) Three centuries of category errors in studies of the neural basis of consciousness and intentionality. Neural Networks 10: 1175-1183.
- Feldman DE (2012) The spike-timing dependence of plasticity. Neuron 75: 555-571.
- Fingelkurts AA, Fingelkurts AA, Neves CF (2012) "Machine" consciousness and "artificial" thought: an operational architectonics model guided approach. Brain Res 1428: 80-92.
- Averbeck BB, Latham PE, Pouget A (2015) Neural correlations, population coding and computation. Nature Rev Neurosci 7: 358-366. [Crossref]
- Marx G, Gilon C (2012) The molecular basis of memory. ACS Chemical Neurosci 3: 633-642.
- Marx G, Gilon C (2018) The molecular basis of neural memory. Part 10. The sins and redemption of neurobiology. J Neurol Neurocrit Care 1: 1-7.
- Marx G, Gilon C (2020) Memory, emotions, language and mind. J Psychology Neuroscience 2: 1-9.
- Ramirez S (2018) Crystallizing a memory. Science 360: 1182-1183.
- Katchalski E (1992) Molecular surface recognition: Determination of geometric fit between proteins and their ligands by correlation techniques. Proc Natl Acad Sci USA 89: 2195-2199.
- Tononi G (2004) An information integration theory of consciousness. BMC Neuroscience 5: 42-69.
- Tononi (2008) Consciousness as integrated information: A provisional manifesto. Biol Bull 15: 216-242.
- Greene B (2000) The Elegant Universe. Vintage Random House, London.
- Becker A (2018) What is Real? John Murray Publishers, UK.
- Bergson H (1912) Matter and Memory. Dover Publications, New York Translation : Allen & Co, 2004. London.
- Brough JB (1975) Husserl on Memory. The Monist 59: 40-62.
- Ryle G (1949) The Concept of Mind. Penguin Books UK.
- Herculano-Houzel S, Manger PR, Kaas JH (2014) Brain scaling in mammalian evolution as a consequence of concerted and mosaic changes in numbers of neurons and average neuronal cell size. Front Neuroanat 8:77.
- Strausfeld N, Hirth F (2015) Introduction to ‘Origin and Evolution of the Nervous System’. Philos Transact B Roy Soc B 370: 20150033.
- Waldrop MM (1992) Compexity: The emerging science at the edge of order and chaos. Viking, New York.
- Johnson S (2001) Emergence: The Connected Lives of Ants, Brains, Cities and Software. Touchstone Books, New York.
- Tadi KK, Alshanski I, Mervinetsky E, Marx G, Petrou P (2017) Oxytocin-monolayer-based impedimetric biosensor for zinc and copper ions. ACS Omega 2: 8770-8778.
- Alshanski I, Blaszkiewicz J, Mervinetsky E, Rademann J, Yitzchaik S (2019) Sulfation patterns of saccharides and heavy metal ion binding. Chem Eur J 25: 1-9.
- Leiserson CE, Thompson NC, Emer JS, Kuszmaul BC, Lampson BW (2020) There's plenty of room at the Top: What will drive computer performance after Moore's law? Science 368: 1079.